КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Популяция как элементарная эволюирующая единица
|
|
|
|
Биоценозы, как правило, не имеют сплошного распространения. Они распределяются по биотопам с соответствующими почвенно-климатическими условиями. Соответственно этому и особи определенного вида организмов не имеют сплошного распространения. Входя в состав определенных биогеоценозов, они распределяются группами по подходящим биотопам, образуя в каждом из них более или менее обособленное население — популяцию особей данного вида организмов. Величина и обособленность популяций могут быть весьма различными и это зависит как от пространственных соотношений (большой и однородный лес или маленькая роща, река, озеро или пруд, безграничная степь, луг в пойме реки или лужайка в лесу, большое заболоченное пространство или отдельное болотце и т. п.), так и подвижности самого организма (способ рассеивания семян у растений, активное перемещение у животных). Во всяком случае жизнь и размножение особей данного вида организмов протекает в популяциях разной величины и разной обособленности, входящих в состав определенных биогеоценозов.
Эволюция определяется борьбой за существование в условиях сложных взаимоотношений между элементами биогеоценоза. Эти соотношения не являются постоянными. Кроме случайных и периодических колебаний, можно установить также наличие исторических изменений состава целых биогеоценозов. Наиболее существенным, основным выражением исторического преобразования живых систем (включая биоценозы) является эволюция морфофизиологической организации особей как представителей определенного вида организмов. Эта эволюция протекает только в ряду поколений, т. е. в целых линиях особей, совместно составляющих данную популяцию во временной преемственности ее структуры. Наиболее типично протекает эволюция в популяциях, состоящих из свободно меж собой скрещивающихся особей. Только при скрещивании происходит перекомбинирование наследственных свойств как важнейший процесс перестройки наследственной основы организации.
|
|
|
Возможность свободного скрещивания фактически ограничивается главным образом лишь особями одной популяции, хотя обычно возможен и более или менее свободный обмен мигрантами между разными популяциями одного и того же вида. Как уже сказано, степень обособленности между популяциями может быть весьма различной и это зависит не только от территориальной их близости, но и от степени подвижности и средств расселения данного вида организмов. Соревнование особей и естественный отбор протекают также в основном внутри популяции и если биогеоценоз является ареной первичных эволюционных преобразований, то входящая в его состав популяция данного вида организмов является наименьшей эволюирующей единицей.
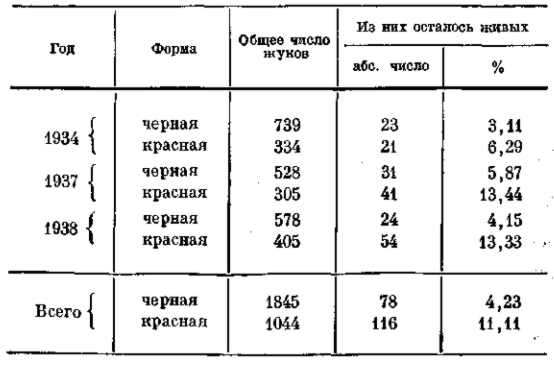
До тех пор, пока вид еще не распался на вполне изолированные новые единицы, пока не прекратились обмен мигрантами и внутривидовые скрещивания, он изменяется в процессе эволюции как более или менее сложная, но все же целостная система. При этом руководящее значение имеют сложные взаимоотношения организмов с внешней средой, названные Дарвином борьбой за существование. Эта борьба связана с гибелью (элиминацией) одних, как правило, менее приспособленных, особей и естественным отбором других, имеющих в среднем известные преимущества в данных условиях внешней среды. В результате этого одни формы исчезают из популяции, а другие получают в ней все большее распространение. Состав популяции при этом меняется. В редких случаях удается непосредственно установить изменение численных соотношений различных типов особей, входящих в состав популяции. Так, например, у божьих коровок (Coccinellidae) популяции одного и того же вида состоят из особей различной окраски. Географически различные популяции содержат те же типы, но в различном численном соотношении (Harmonia axyridis, рис. 10, Dobzhansky Т., 1933, 1937). Различное направление естественного отбора ведет к изменению этого соотношения даже в одной и той же изолированной популяции по сезонам. Весной у Adalia bipunctata (рис. 18, табл. 2) преобладает «красная форма», осенью— «черная», которая летом, очевидно, размножается с большей интенсивностью, а зимой гибнет в большем числе (Timofeeff-Rossovsky, 1940).
|
|
|
Географическое изменение численного соотношения между альтернативными типами в популяциях наземных моллюсков рода Partula, населяющих различные долины островов Таити и Муреа, установлено Крэмптоном (Crampton, 1932). Популяции даже соседних долин различаются по комбинациям признаков, охватывающих окраску, величину и форму раковины, правое и левое вращение раковины моллюска.
Гораздо чаще популяции имеют фенотипически однородный характер. Тогда географическая изменяемость представляется первоначально непрерывной по всем признакам. Дж. Гексли (J. Huxley, 1940) говорит в этом случае о «градиенте» признаков (Charakter-gradients, clines). Несмотря на непрерывность связи, географически наиболее удаленные представители такого вида могут быть настолько различны, что не только по морфологическим признакам, но и физиологически (по нескрещиваемости) заслуживают выделения в самостоятельные виды. Много фактов такого рода собрано у Ренша (Rensch, 1929, 1939). Нас, однако, занимает здесь не самая проблема видообразования, а лишь процесс возникновения стойких различий.
Хорошим примером непрерывной географической изменяемости может служить из растений обыкновенный сон-прострел (Anemone Pulsatilla, Zimmermann, 1938). Он дает непрерывный ряд форм, закономерно изменяющихся в направлении с запада на восток Европы. На западе он обладает тонко рассеченными и разбросанными листьями и поникшими цветами. На востоке у него более грубо рассеченные, стоячие листья и стоячие цветы. Эти различия имеют наследственный и вместе с тем приспособительный характер — они связаны с обилием дождей на западе и засушливостью в восточной Европе. Положение листьев способствует стеканию дождевой воды к корневой системе у восточных форм и, наоборот, разбрызгиванию воды у западных. Поникшее положение цветов предохраняет от смывания пыльцы дождевой водой. Степень рассеченности листьев связана с различной интенсивностью транспирации. Мало рассеченные листья восточной формы испаряют значительно меньше воды, чем богато расчлененные листья западной формы (что доказано прямым измерением транспирации в одинаковых условиях опыта). Из растений можно еще привести подобный пример переходов между горной формой люцерны Medicago glutinosa и степной формой Medicago jalcata. На горных лугах Балкарии и Карачая встречается типичная М. glutinosa с опушенными, спирально закрученными плодами (бобами). При переходе на нижележащие луга опушение теряется и плод раскручивается. Medicago glutinosa постепенно переходит в типичные формы М. jalcata, которые в степи, на границе гор, обладают прямыми и голыми плодами (Синская, 1938).
|
|
|
ТАБЛИЦА 2. Число переживающих зиму красных и черных божьих коровок Adalia Bipunctata по результатам подсчета на местах зимовки в предместье Берлина
Примером непрерывной географической изменяемости могут служить улитки Cerion на Багамских островах. На острове Нью-Провидаис они постепенно и закономерно изменяются при переходе с запада на восток. Западные формы — белые и ребристые; восточные — слабо ребристые или почти гладкие и пигментированные. Все они связаны совершенно непрерывными переходами, дающими полный ряд промежуточных форм между обоими крайними типами. Изменения, по-видимому, связаны с различиями в климате — на востоке более дождливом, на западе — засушливом. Более толстая, белая раковина западных форм, очевидно, лучше защищает тело от чрезмерного нагревания на солнце (L. Plate, 1907).
Постепенный ход изменений установлен В. Алпатовым (1927) для домашней пчелы. При переходе с севера на юг можно отметить уменьшение общих размеров, удлинение конечностей и удлинение хоботка. Последнее связано, очевидно, с использованием других цветов, обладающих более глубокими нектарниками. Жук-жужелица Discoptera komarovi дает при биометрической обработке картину непрерывного ряда изменений в направлении с востока Туркестана на запад до Каспия. Закономерно меняется целый ряд признаков. Приспособление к жизни в барханных песках ведет ко все большей специализации западных популяций. Очевидно, эволюция этого жука сопровождалась его постепенным распространением. Восточные популяции D. komarovi филогенетически старше и стоят по своей организации ближе к исходным туркестанским формам. Прикаспийские популяции наиболее молоды, но и наиболее изменены (К. Арнольди, 1939).
|
|
|
Аналогичные исследования, сопровождавшиеся генетическим анализом, были проведены в большом масштабе Р. Гольдшмидтом (R. Goldschmidt, 1927, 1935) над непарным шелкопрядом (Porthetria dispar). Этот вид обладает широким распространением в Европе и Азии и распадается на много географических рас, связанных между собой всеми переходами. Они различаются по общей величине, по пигментации и другим признакам. Они различаются и по своему жизненному циклу, и здесь ясно видна приспособительность расовых различий по отношению к местным условиям климата. Северные и горные популяции, соответственно более краткому периоду вегетации, обладают более быстрым эмбриональным развитием и более коротким периодом личиночного развития, чем южные расы. Здесь, как видно, преобладают физиологические различия.
В других случаях именно физиологические различия выступают на первый план, и географические расы морфологически еще неразличимы. Так, у Drosophila funebris установлено существование географических рас, обнаруживающих приспособительные различия в стойкости по отношению к крайним температурам и в температурном оптимуме. Восточная раса, населяющая СССР, весьма устойчива как к высоким, так и к низким температурам. Северо-западная раса довольно устойчива к низким температурам, в то время как юго-западная раса, распространенная по средиземноморским странам, весьма устойчива по отношению к высоким температурам (Timofeeff-Ressovsky, 1935a). Сходные физиологические различия (по плодовитости в различных условиях температуры) были установлены для различных популяций Drosophila pseudoobscura (Dobzhansky, 1937). Здесь мы имеем, несомненно, лишь самые начальные этапы дифференцировки единого вида на местные географические или экологические формы.
Мы не ставим себе задачей последовательный анализ процессов видообразования. Это делалось многими современными авторами (В. Л. Комаров, 1940; Rensch, 1939; Dobzhansky, 1937; Сборник под ред. J. Huxley — The new systematica, 1940, и др.). Обратим, однако, внимание на следующие факты. Генетический анализ указанных межрасовых различий показал,— во всех случаях, когда такой анализ был возможен, — наличие наследственных изменений (хотя наряду с этим могли быть и модификационные изменения, например, у той же Anemone Pulsatilla). Эти различия имеют дискретный характер мутационных изменений. При скрещивании различных рас они показывают картины менделевского расщепления.
Однако в подавляющем ряде случаев различия касаются огромного числа «генов», т. е. представляют результат комбинирования весьма многочисленых мутаций. Гибриды первого поколения обычно довольно однородны, а во втором поколении обнаруживается значительное увеличение изменчивости по всем признакам. Это указывает на подавляющее значение многочисленных малых мутаций при формировании географических (и экологических) рас.
Уже простые различия в окраске божьих коровок Harmonia ахуridis (от желтых с черными пятнами до черных с желтыми пятнами) определяются изменениями в трех генах (Tan and Li, 1934). По Семнеру (Sumner, 1920, 1932), географические изменения грызунов Peromyscus определяются изменениями очень многих генов, что доказывается сильным возрастанием изменчивости во втором поколении гибридов. Гольдшмидт (R. Goldschmidt, 1927–1935) находит, что различия географических рас непарного шелкопряда Porthetria dispar простираются на «силу» половых факторов, на окраску гусениц, на величину и окраску взрослых бабочек, на длительность зимнего покоя яиц, на скорость развития и другие признаки. Для каждого признака можно установить влияние целого ряда «генов», одни из которых вызывают значительный эффект, а другие модифицируют его в сторону усиления или ослабления.
Многочисленные мелкие наследственные различия установлены и Бауром (Е. Ваш, 1924) при анализе различных природных популяций львиного зева (Antirrhinum). Наконец, и сравнение местных популяций и рас различных видов дрозофилы показало существование многочисленных различий, основанных, главным образом, на различной концентрации одних и тех же генов. Встречаются и более крупные различия, связанные со стерильностью гибридов между такими популяциями (расы А и В у Drosophila pseudoobscura).
В некоторых случаях различия популяций и рас имеют как будто иной характер и определяются изменением одного только гена. В таких случаях говорят о распространении одной простой мутации. Известным примером такого распространения являются мела-нистические хомяки (Cricetus cricetus), изученные сначала зоогеографом Лепехиным, а затем рядом других исследователей; в новейшее время их распространение изучали С. В. Кириков (1934) и С. М. Гершензон (1946). Они концентрируются у нас главным образом в двух очагах — в Башкирии и на Украине. В обоих случаях их ареал ограничивается более влажными зонами лесостепи.
В Западной Германии описана также простая рецессивная мутация полевки (Microtus arvalis), отличающаяся измененным строением коренных зубов. Ее концентрация достигает в Шлезвиг-Гольштейн, очевидном очаге ее распространения, 85% всех особей. По направлению на юг относительная численность мутантов быстро падает (Zimmermann, 1935).
У божьей коровки Epilachna chrysomelina (Coccinellidae) известна полудоминантная мутация Elaterii, которая характеризуется определенным изменением рисунка. Она образовала в части средиземноморской области (Сев. Италия, Южн. Франция, Корсика, Сардиния) хорошо выраженную географическую расу. Генетически она составляется из одной большой мутации и ряда малых.
Это последнее представляет, очевидно, почти общее правило, относящееся ко всем случаям распространения «отдельных», более значительных мутаций.
Исследования межвидовых гибридов хлопчатника (Харланд, 1937) показали, что этим же путем могут устанавливаться и видовые различия, в которых всегда участвуют многочисленные малые наследственные изменения, хотя ведущее значение и могла иметь всвое время единичная мутация. В огромном большинстве случаев ясно выраженные географические расы отличаются весьма значительным числом генных мутаций, большинство которых связано с незначительными количественными изменениями во многих морфологических и физиологических признаках. Нередко можно установить и приспособительный характер различий, п это указывает тогда на ведущую роль естественного отбора в их накоплении. Вместе с тем ясно, что естественный отбор действует всегда на фоне множества мелких различий, имеющихся между особями любой популяции. Концентрация определенных изменений (мутаций) в определенных популяциях и притом не всегда явно приспособительных; изменения этой концентрации по сезонам, по периодам колебания общей численности; существование известных различий в накоплении мутаций в больших и малых популяциях,—все это указывает на существование довольно сложного комплекса факторов, определяющих динамику исторического преобразования популяции. В этом отношении имеют значение прежде всего, конечно, борьба за существование и естественный отбор, но кроме того — также комбинирование в процессах скрещивания и изоляция, т. е. ограничение этого скрещивания. Сверх того мы должны отметить значение и некоторых менее явных процессов развития системы регуляторных механизмов формообразования и накопления мутаций в скрытом виде, что приводит к внешне незаметной перестройке генотипа. Наконец, мы подчеркнем и значение индивидуальной приспособляемости, основанной на широком развитии системы реакционных механизмов.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 443; Нарушение авторских прав?; Мы поможем в написании вашей работы!