КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Формы борьбы за существование 2 страница
Очевидно, среди этих рыб преобладало значение активного соревнования, которое и привело к богатому экологическому расхождению. Элиминация имела здесь избирательный характер.
С другой сторны, в озерах Альберт и Рудольф имеются большие активные хищники: «нильский окунь» и «рыба тигр». Соответственно этому другие рыбы, которыми эти хищники кормятся, не дали заметной радиации форм. Очевидно, их прямая истребляемость имеет почти неизбирательный характер. Борьба за существование оказывается в значительной мере пассивной.
Отмечается также удивительный консерватизм двудышащей рыбы Protopterus, которая ведет весьма малоактивный образ жизни и в то же время не дала заметного видообразования. Другие примеры будут разобраны далее. Здесь мы должны только подчеркнуть, что избирательная элиминация, а следовательно и отбор, захватывают, конечно, организм в его морфофизиологическом выражении, т. е. по фенотипу. Непроявляющаяся мутация не может иметь избирательного значения. Поэтому все формы скрытого мутирования, связанного с рецессивностью мутации в гетерозиготном диплоидном, а тем более тетраплоидном организме, или с высокой регулярторной способностью организма, не являются непосредственным объектом отбора. Однако они могут иметь косвенное значение, определяя стойкость организма в известных условиях и его пластичность, при их изменении. Об этом будет речь в дальнейшем изложении.
Естественный отбор является процессом, обратным по отношению к элиминации, однако только по отношению к избирательной элиминации, да и то с известными оговорками. Поэтому острота элиминации ни в какой мере не определяет интенсивности естественного отбора. Это создает трудности для учета реальных интенсивностей отбора в природных условиях. Об этой интенсивности можно иметь суждение лишь по реальной скорости распространения известной формы в известных условиях существования. Но указанная скорость определяется и другими факторами, которые нам еще предстоит рассмотреть.
Математическая теория естественного отбора, разработанная Фишером (Fisher R. А., 1930), Холденом (Haldane J. S., 1924), Райтом (Wright S., 1931) и др., исходит из теоретических предпосылок и по необходимости крайне упрощает стоящую перед нею задачу. Однако и при таком упрощении задачи большинство вопросов требует для своего разрешения чрезвычайного напряжения математических средств. Мы можем в дальнейшем разборе принять во внимание лишь некоторые основные выводы этих авторов, касающиеся вопросов соотношения мутабильности, отбора и изоляции в процессах изменения генетической структуры популяций. Здесь же мы отметим лишь некоторые качественные характеристики элиминации и отбора, на которые пока обращалось недостаточно внимания, хотя они имеют большое значение.
Нередко говорят об уничтожающей роли естественного отбора вообще, или об отрицательном отборе в специальном случае уничтожения неприспособленных форм. Во всех случаях уничтожения отдельных особей или целых групп особей я предпочитаю говорить об элиминации. При элиминации оформленных таксономических групп (рас, подвидов, видов и высших таксономических подразделений) можно говорить о вымирании. Термин «отрицательный отбор» отличается двусмысленностью, так как в применении к известным признакам он получает противоположное значение: элиминация ветром летающих насекомых небольших океанических островов («отрицательный отбор») приводит к переживанию, т. е. положительному отбору нелетающих особей. В данном случае также нередко говорят, что редукция крыльев есть результат «отрицательного отбора» (по недоразвитым крыльям).
Во всех случаях уничтожения отдельных особей или местных популяций мы будем говорить об их элиминации.
Межгрупповое соревнование, связанное с элиминацией целых групп особей (популяций, подвидов, видов и др.), ведет к переживанию одних форм (отбор) и уничтожению других (вымирание). В этом случае отбор происходит между уже существующими формами. Он не имеет здесь явного творческого значения. Говорят об отсеивающей роли отбора (сравнение с решетом) или об его уничтожающей роли, так как в результате его действия число групп и, следовательно, многообразие органических форм уменьшается. Таково значение отбора при межгрупповом соревновании. Сюда относится дарвиновское вымирание промежуточных форм, ведущее к таксономическому расхождению организмов. Так как группа, уничтожаемая или вытесняемая в одной местности, может получить преобладание в другой, то межгрупповое соревнование (групповая элиминация) связано с распределяющей ролью отбора. Однако территориальное и экологическое распределение организмов связано с их приспособленностью к известным условиям существования. В этом случае мы имеем дело с положительным фактором эволюции флор и фаун, т. е. именно С деятельностью естественного отбора. Однако последний оперирует с готовыми формами и, следовательно, не имеет здесь творческого значения.
Определенно творческое значение имеет лишь индивидуальный естественный отбор, основанный на соревновании особей внутри известной группы (популяции, расы, подвида, вида), т. е. на индивидуальной элиминации. Он действует всегда на фоне подвижного материала, непрерывно меняющегося в процессах мутирования и комбинирования. Последнее определяется скрещиванием. Комбинирование и даже само мутирование идет под постоянным контролем естественного отбора. Благоприятные комбинации не возникают случайно — они создаются под направляющим действием естественного отбора. Именно эта роль естественного отбора, творящего новые формы, и составляет предмет нашего рассмотрения (в остальном см. И. Шмальгаузен, 19386, 1939а).
Творческая роль естественного отбора проявляется также в различных формах в зависимости от данных соотношений между организмом и факторами окружающей среды.
При изменении условий внешней среды приспособленность организма к этой среде, естественно, теряется, и его элиминация возрастает. При наличии таких индивидуальных уклонений, которые повышают приспособленность организмов к новым условиям существования, они будут переживать в большем числе и в результате размножения получать все большее распространение в популяции. Особи, представляющие прежнюю норму, окажутся в худшем положении и будут гибнуть в относительно большем числе. Популяция начнет изменяться приспособительно к новым условиям существования и таким образом установится новая норма на месте старой.
Это — обычная дарвиновская борьба за существование, связанная с прогрессивной эволюцией на основе элиминации прежней нормы. Естественный отбор выступает здесь в роли ведущего фактора эволюции, изменяющего старую и создающего новую, в данном случае более приспособленную, норму.
Если же факторы внешней среды получили довольно устойчивый характер и организм уже вполне приспособлен к данным условиям существования, то преимущества в борьбе за существование будут на стороне установившейся нормы, а уклонения, нарушающие приспособленность организма, начнут гибнуть в гораздо большем числе.
Несмотря на наличие дарвиновской борьбы за существование, заметных изменений в организации не произойдет. Наоборот, естественный отбор будет поддерживать существование данной нормальной организации (хотя и будут происходить изменения в процессах индивидуального развития, приводящие к увеличению его устойчивости). Отбор выступает здесь в роли стабилизирующего фактора, действующего на основании э л и м и н а-ц и и всех уклонений от нормы.
Таким образом, две формы элиминации определяют два различных выражения естественного отбора — его ведущую роль (меняющую норму) и его стабилизирующее значение (укрепляющее установившуюся норму). Если бы не указанная двусмысленность в понимании «отрицательного» отбора — 1) как элиминации менее приспособленных особей вообще и 2) как отбора на уменьшение известных признаков, то, принимая второе определение, можно было бы различать следующие основные формы отбора:
I. Отбор положительный (ведущая форма), идущий в направлении нарастания и усложнения признака. В результате происходит прогрессивное развитие данной части.
П. Отбор отрицательный (ведущая форма), идущий в сторону уменьшения и упрощения признака. В результате происходит регрессивное развитие данной части.
III. Отбор нейтральный (стабилизирующая форма), поддерживающий установившуюся нормальную величину и строение признака. В результате происходит стабилизация формообразования, т. е. автономизация онтогенеза и развитие его регулятор-ных механизмов.
6. ФОРМЫ ОТБОРА
Естественный отбор меняет не только свое направление, но и свои формы в зависимости от изменений условий и форм борьбы за существование. В борьбе за существование находят свое многообразное выражение сложнейшие взаимодействия между организмом и окружающей его неорганической и органической средой (включая и взаимозависимости между особями одного и того же вида организмов). Поэтому как борьба за существование, так и естественный отбор изменяются в своей интенсивности и в своей качественной характеристике при изменении любых факторов внешней среды, так же как и при изменении строения, функций и нормы реакций самого организма.
Мы здесь не ставим своей задачей анализ всех возможных соотношений, но фиксируем наше внимание на значении характера самих изменений (неуклонных, т. е. направленных, периодических или колеблющихся) однородной или разнородной среды, а также на изменчивости лабильных и стабильных организмов, обнаруживающих в своем формообразовании различную степень зависимости от факторов внешней среды.
Ведущая (движущая) форма отбора реализуется на основе селекционного преимущества (в измененных условиях внешней среды) определенных уклонений организации перед нормой, установившейся в прежних условиях существования. Она связана с частичной элиминацией прежней нормы и установлением новой. Стабилизирующая форма отбора осуществляется на основе селекционного преимущества (в известных, особенно в колеблющихся, условиях) нормальной организации перед уклонениями от нормы. Она связана с элиминацией большинства уклонений и с выработкой более устойчивых механизмов нормального формообразования.
А. ВЕДУЩАЯ ФОРМА ОТБОРА
а. Стабильный организм в однородной неуклонно меняющейся среде. Наиболее простыми являются изменения соотношений между однородной средой, меняющейся в одном определенном направлении, и изменчивым организмом со стабильным (т. е. независимым от изменений внешних факторов) формообразованием. Это — классическая форма естественного отбора, к которой относится и большинство его доказательств. Его действие можно пояснить следующими схемами (рис. 9, А, В).
Первый рисунок (А) показывает типичную вариационную кривую (1), характеризующую известный признак всех особей некоторой популяции определенного вида организмов. Вторая кривая (2) характеризует изменчивость того же признака «нормальных» особей, переживающих и оставляющих потомство. Промежуток между обеими вариационными кривыми представляет зону вариаций, элиминируемых в борьбе за существование.
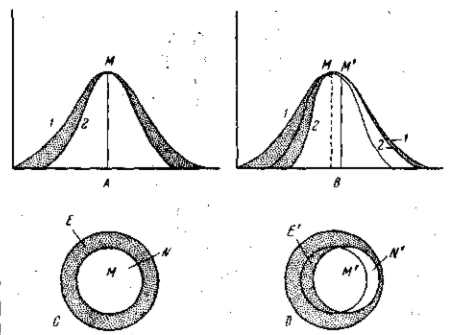
Рис. 9. Отбор в однородных условиях внешней среды. А — отбор при установившихся условиях (стабилизирующая форма отбора); 1 — вариационная кривая всех особей; 2 — вариационная кривая особей, выживающих и оставляющих потомство («норма»). Заштрихована область вариантов, элиминируемых при данных условиях. В —отбор при изменении этих условий в определенном направлении (движущая форма отбора): 1 — вариационная кривая всех особей; 2 — вариационная кривая особей, выживающих и оставляющих потомство в новых условиях (новая «норма»); С, D — другой план изображения механизма действия стабилизирующей и движущей форм отбора
При изменении внешней среды в определенном направлении, нарушающем приспособленность организма по данному признаку, некоторые особи прежней нормы окажутся менее приспособленными, а некоторые варианты, ранее устранявшиеся, могут приобрести известные преимущества. В результате этого, с одной стороны вариационной кривой (рис. В) произойдет сокращение численности переживающей нормы и соответственное расширение зоны элиминируемых вариаций (—), а с другой, благоприят-ствуемой, стороны произойдет расширение нормы за счет зоны вариаций, которые ранее элиминировались (+). Кривая переживающих особей примет асимметричный вид и средняя сдвинется в благоприятствуемую сторону (мода может вначале оставаться на месте и в своем движении отстает от средней тем более, чем резче меняется среда и чем жестче элиминация). Накопление вариаций, а следовательно — и мутаций, которые ранее элиминировались, ведет к некоторому нарушению регуляторного аппарата («генного баланса» и всей системы морфогенетических корреляций вообще), а это, в свою очередь, способствует проявлению мутаций, которые были погашены в своем выражении (вскрытие резервов, см. Н-5), и дальнейшему повышению мутабильности. Поэтому вариационная кривая расширяется, и тогда начинается ее движение в благоприятствуемом направлении за пределы вариантов, наблюдавшихся ранее (рис. У). Этот процесс идет медленнее, чем первая фаза эволюции,—сдвиг средней за счет наличных уже вариантов и мобилизации скрытых резервов внутривидовой изменчивости. Однако в благоприятных условиях эволюция приобретает затем характер самоускоряющегося движения и может идти все же с очень значительной скоростью (см. раздел IV). То же самое можно иллюстрировать и диаграммами, представляющими как бы поперечное сечение трехмерных вариационных кривых (рис. 9, С, D).
Большинство прямых и косвенных доказательств существования естественного отбора касается именно этой наиболее простой и наглядной его формы. Из косвенных доказательств можно сослаться на явления покровительственной окраски и мимикрии, а также на другие средства пассивной защиты. Совершенно ясным примером того же действия естественного отбора может служить редукция крыльев у островных насекомых или редукция органов зрения у роющих позвоночных.
Из прямых доказательств ведущей роли естественного отбора у стабильных организмов можно упомянуть наблюдения Уэлдона (Weldon, 1898) над крабами Carcinus maenas в гавани Плимута, где, в связи с постройкой мола и замутнением воды, произошло сужение головогрудного щита (очевидно, это лишь первая фаза процесса — сдвиг средней в пределах имевшихся уже вариаций). Сюда же относятся наблюдения Гаррисона (Harrison, 1920) над изменением окраски бабочек Oporabia autumnata в сторону посвет-ления при замене соснового леса березовым. Наконец, из новейших исследований такого рода следует упомянуть наблюдения Дубовского (1941) над приспособлениями пресноводных и морских ракушковых рачков (Ostracoda), сопровождавшиеся экспериментальными исследованиями значения некоторых адаптации (средств прикрепления) и отбора в условиях текучей воды.
б. Лабильные организмы в однородной неуклонно меняющейся среде. В случае эволюции лабильного организма наша простая схема полностью сохраняет свое значение, но усложняется лишь за счет наличия модификационных изменений, прикрывающих мутационные изменения и потому затрудняющих их учет. Ведущая форма отбора комбинируется здесь со стабилизирующей, о которой речь будет идти в дальнейшем. Все же направленность эволюции бывает ясно выражена, и движущая роль отбора не вызывает сомнений. Зона элиминируемых вариаций составляется в этом случае, кроме мутаций, также из модификаций (из морфозов и адаптивных реакций на случайные, кратковременные изменения в факторах среды, т. е. из реакций, оказавшихся «неудачными» или «ошибочными»). Отбор положительных наследственных уклонений происходит на фоне более или менее значительных и более или менее совершенных индивидуальных приспособлений (модификаций).
Многие географические изменения, отличающиеся своей непрерывностью, характеризуются именно теснейшим соединением мутационных и модификационных изменений, идущих в одном и том же направлении.
Так, например, чистые линии льна, высеянные в разных широтах, дают фенотипические изменения, сходные с географическими формами данных мест, отличающиеся, однако, и по своему генотипу. Северный лен-долгунец отличается большей высотой и меньшим числом стеблей и коробочек, чем южный лен-кудряш. Это — наследственные отличия. Однако на юге лен-долгунец становится ниже, а число стеблей и коробочек увеличивается. Наоборот, на севере лен-кудряш до известной степени приближается к долгунцу. Сон-прострел (Anemone Pulsatilla) обладает в своей западной расе разбросанными и мелко рассеченными листьями, а также поникшими цветами. У восточной расы листья приподняты, более грубо рассечены и цветы стоячие. Это — наследственные различия. Их гибриды менделируют. В их потомстве происходит расщепление по форме листьев в отношении 25% особей, обладающих листьями с широкой верхушкой, 25 % — с узкой и 50 % — со средней верхушкой, как у гибридов. Однако эти наследственные различия перекрываются сходными модификационными изменениями, которые могут быть получены в опыте при изменении интенсивности освещения (соответственно меньшей облачности восточной Европы) !.
Северные млекопитающие и птицы отличаются более крупными размерами от южных представителей той же группы (правило Бергманна). Эти отличия — наследственные, и они могли установиться в результате естественного отбора на более экономичный обмен и меньшую потерю тепла у северных форм. Однако частично эти различия могут иметь характер модификаций, как показывают некоторые эксперименты. Точно так же более короткие уши и хвосты северных грызунов и некоторых других млекопитающих (правило Аллена) являются наследственными свойствами, развившимися в процессе естественного отбора на меньшую потерю тепла (через уши и хвост, обладающие слабым шерстяным покровом и служащие для теплорегуляции). То же самое касается густоты общего шерстяного покрова, длины волос и их толщины у северных млекопитающих по сравнению с южными расами и подвидами. Однако и здесь наследственные изменения перекрываются сходными модификационными (как видно из опытов Семнера).
в. Стабильные организмы в разнообразной неуклонно меняющейся среде. Естественный отбор в однородной среде, конечно, никогда не осуществляется в чистом виде. Наиболее единообразны разве лишь условия существования паразитов, частично — глубоководных или пещерных животных, животных, зарывающихся в более глубокие слои почвы или в грунт более глубоких вод.
За этими лишь исключениями, факторы среды меняются в зависимости от места пребывания, от времени года и даже суточных или случайных колебаний освещения, температуры, влажности и т. п. Внешняя среда не бывает однородной ни в пространстве, ни во времени, ни по своим физическим факторам, ни по биологическим (пища, враги, паразиты).
В этом случае наша схема ведущего отбора усложняется в том отношении, что никакая «норма» не может быть одинаково приспособленной и ко всем местным условиям среды и ко всем ее как случайным, так и периодическим изменениям.
Поэтому само понятие «нормы» становится несколько неопределенным. Мы все же будем понимать под «нормой» просто всю сумму особей, которые фактически переживают в конкретных (разнообразных) условиях существования и оставляют после себя потомство. Это, однако, лишь условная средняя норма, которая фактически непрерывно меняется.
Кроме зоны безусловно элиминируемых уклонений, теперь приходится учитывать существование зоны условно благоприятных уклонений, которые в некоторых частных условиях существования получают известные преимущества перед средней нормой. В некоторых местностях, в известные сезоны или при случайных уклонениях в факторах среды переживают и оставляют потомство не представители средней нормы, а эти условно благоприятные варианты. В других местностях, при других уклонениях эти варианты элиминируются, но зато переживают другие варианты. При дружной весне все преимущества будут на стороне рано и быстро развивающихся растений и рано вылупляющихся личинок насекомых (обладающих низким температурным порогом реактивности). Однако в следующий год вернувшиеся весной морозы могут убить их потомство, и тогда все преимущества будут на стороне поздно и медленно развивающихся особей. Отбор меняет свое направление, и это поддерживает весьма значительную гетерогенность популяций растений и животных, живущих в разнообразной и непостоянной среде.
В случае существования ясно очерченных локальных (экологических) условий гетерогенность популяций принимает формы более или менее ярко выраженного генетического полиморфизма (гетероморфизма). Так должен был развиться гетероморфизм богомолов (Mantis religiosa), обладающих зелеными и буро-желтыми формами, которые прекрасно защищены в зеленой листве, на бурых стеблях кустарников и на желтой, высыхающей летом траве. Эти основные формы имеют наследственный характер и непрерывно выщепляются в чистом виде у родителей любой окраски (М. М. Беляев, 1946).

Рис. 10. Основные типы окраски у божьей коровки Harmonia axyriiis. а — var succinea, b — var frigida, с — var 19-signata, d — var aulica, e — var axyridis, f — var spectabilis, g — var conspicua. По Ф. Добжанскому, 1937
Таково же значение гетероморфизма мимикрирующих самок африканских папилионид (Papilio polytes и P. dardanus), которые находят свою защиту в сообществе различных иммунных видов (относящихся к родам Amauris и Danais). Каждая форма имеет свою самостоятельную линию эволюции, как это показывает факт образования у P. dardanus в Ю. Африке ряда локальных географических форм, копирующих местные подвиды Amauris echeria. Между тем это один вид, непрерывно выщепляющий различные, характерные для данной местности, формы самок.
Холден указывает, что в некоторых случаях гетероморфизм может быть очень устойчивым. Так, в современных популяциях садовых улиток Cepea hortensis и С. nemoralis встречаются гладкие (однотонные) и полосатые формы. Последние несколько преобладают. В отложениях неолита типы этих же улиток найдены почти в современных пропорциях. В плейстоцене оба типа были приблизительно равночисленными. Следовательно, если и имеется определенный сдвиг, он совершался очень медленно (Lang, 1912).
В других случаях нетрудно установить не только географические, но и сезонные изменения направления отбора. У божьих коровок имеется множество вариантов, выражающихся в окраске (рис. 10). Некоторые виды оказываются резко гетероморфными. Так, Ф. Добжанский изучал географические расы Harmonia axyridis, отличающиеся необыкновенным разнообразием в положенной распространении пятен. Почти во всех расах повторяются одни\ и те же красочные типы, однако в различных пропорциях. При расселении этого вида в одних местностях получали условное преимущество одни формы, в других — другие. Тимофееву-Ресовскому удалось показать даже изменение направления естественного отбора по сезонам. У божьей коровки Adalia bipunctata в осенней популяции преобладает «черная» форма. Однако, очевидно в силу большей ее смертности во время зимовки, весной преобладает «красная» форма. По Лукину (1939), у клопа Pyrrhocoris apterus наблюдаются и локальная и сезонная изменчивость размеров тела.
г. Лабильные организмы в разнообразной неуклонно меняющейся среде. При изменении лабильного организма в разнообразной среде картина эволюции усложняется наложением модификации на существующий уже генетический полиморфизм.
Географические изменения высших растений происходят главным образом по этому типу комбинирования естественного отбора в гетероморфных популяциях (с обособлением «экотипов») и прямого приспособления (выражаемого в «экофенах», по терминологии Турессона). Однако эволюция птиц и млекопитающих также идет в значительной мере по этому пути.
Чем выше значение адаптивных модификаций, тем менее заметную роль играет (на данном отрезке времени) естественный отбор. Руководящая роль переходит тогда на время к адаптивной модификации.
Б. СТАБИЛИЗИРУЮЩАЯ ФОРМА ОТБОРА
Стабилизирующая форма отбора ведет, через элиминацию всех случайных уклонений, к повышению устойчивости существующей уже или устанавливающейся нормы. Б этом случае имеется в виду, что все организмы до известной степени лабильны, т. е. так или иначе, по меньшей мере на ранних стадиях развития, реагируют на изменения в факторах внешней среды, причем эти реакции далеко не всегда оказываются благоприятными (особенно если эти реакции были ответом на случайные и кратковременные изменения). При изменении факторов внешней среды, а также в процессе мутирования, устойчивость формообразовательных систем нарушается (они лабилизируются). В процессе эволюции они в той или иной мере восстанавливаются благодаря непрерывной деятельности стабилизирующего отбора. Результатом этой деятельности является стабилизация форм вследствие создания регу-ляторного аппарата, защищающего нормальное формообразование от возможных нарушений со стороны случайных уклонений в факторах внешней среды, а также и со стороны небольших уклонений во внутренних факторах (т. е. мутаций). Естественные отбор идет в этом случае на основе селекционного преимущества нормы (в частности — новой нормы) перед любыми от нее уклонениями.
а. Стабилизирующий отбор в однородных условиях. В природе лишь редко встречаются виды организмов, живущие во вполне однородных условиях внешней среды. Это касается в известной степени лишь специализированных паразитов, глубоководных животных и других подобных организмов, живущих в весьма ограниченной среде.
Поэтому мы ограничимся вначале лишь общим рассмотрением механизма стабилизирующего отбора в этих наиболее простых условиях, реально всегда осложняемых существованием локальных или сезонных (б), а также случайных (в) изменений в факторах внешней среды. Известные наблюдения Бумпуса над воробьями, погибшими в результате бури (Bumpus, 1899), ясно показывают наличие стабилизирующего отбора в природе. Биометрические исследования погибших воробьев показали, что большинство из них представляло в том или ином отношении уклонения от обычной нормы. Ясно, что именно нормальная организация оказалась наиболее устойчивой при несколько большем напряжении жизненных сил, которого потребовала внезапно налетевшая буря.
Роль стабилизирующего отбора, охраняющего норму, видна в особенности ясно при более детальном генетическом анализе процессов, постоянно протекающих в хорошо изученном материале опытных культур дрозофилы (Drosophila melanogaster), а также при анализе генетического состава природных популяций той же дрозофилы (то же доказано и для других ее видов).
Процесс мутирования, приводящий обычно к более или менее заметному уклонению от «нормального» фенотипа, находит противодействие в естественной элиминации всех вредных уклонений. Однако малые уклонения, лежащие еще в пределах условной нормы (в гетерозиготном состоянии), т. е. фактически способные созреть и оставить потомство, полностью не устраняются и участвуют через скрещивание в постоянной перестройке генотипа данной популяции. Если вредные (условно вредные) уклонения прямо не элиминируются, то они постоянно обезвреживаются через отбор наиболее жизненных комбинаций. Это показывают многочисленные опыты культивирования различных мутаций дрозофилы в лаборатории, при которых степень выражения вредной мутации непрерывно падала. Что это было результатом отбора (т. е. преимущественного переживания и размножения особей, менее уклоняющихся от нормы), видно на опыте скрещивания разных линий с угасшим выражением этой мутации (например, eyeless или stubbloid). После такого скрещивания выражение мутации по меньшей мере частично восстанавливалось. Это означает, ч\го небольшие наследственные уклонения, подавляющие выражение культивируемой мутации («гены-модификаторы») и отобранные во время ведения изолированных культур обеих линий, при скрещивании большей частью перешли в гетерозиготное состояние (так как в различных линиях в большинстве возникали и отбирались разные модификаторы). В гетерозиготе модификаторы потеряли свое выражение и основная мутация вновь проявлялась. Такие же факты были установлены и при культивировании мутаций бокоплавов (Gammarus chevreuxi).
При устранении специфического выражения вредной мутации мы имеем восстановление основных выражений «нормального» фенотипа (при измененном генотипе). Это восстановление достигнуто систематической элиминацией всех более резких уклонений от нормы, т. е. стабилизирующим отбором особей наиболее жизнеспособных, или, иными словами, наиболее приближающихся к установившейся норме.
Существование того же самого процесса установлено, однако, и в популяциях дрозофилы. Природные популяции оказываются насыщенными огромным количеством мутаций (Четвериков, 1926; Тимофеев-Ресовский, 1927; Р. Берг и др., 1941, 1944а, б. См. также сводку Н. Дубинин, 1940а). Кроме рецессивных мутаций, в популяциях оказываются и многие доминантные мутации, близкие к нейтральным или даже вредные (Р. Берг), выражение которых подавлено в генотипе данной популяции или значительно снижено их проявление (Гершензон). Популяция сохраняет свой нормальный «дикий» фенотип, несмотря на непрерывное мутирование, сопровождающееся именно накоплением мутации и, следовательно, непрерывной перестройкой ее генотипа.
Это и есть механизм стабилизирующего отбора, непрерывно «охраняющего» норму и восстанавливающего ее при нарушениях, вызываемых отдельными, не слишком вредными или условно «нейтральными» (в гетерозиготном состоянии) мутациями. При погашении выражения мутации несомненно возрастает и индивидуальная стабильность, которая у «нормальной» особи всегда выше, чем у мутанта.
|
|
Дата добавления: 2015-05-06; Просмотров: 464; Нарушение авторских прав?; Мы поможем в написании вашей работы!