КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Формы борьбы за существование 6 страница
|
|
|
|

Рис. 15. Типичное строение крыльев и гальтеров всех возможных комбинаций аллеломорфов vestigial у Drosophila melanogaster. (No-wing, vestigial, noched, nicked и «дикая» норма), Цифрами обозначен процент краевых вырезок и относительный количественный объеме для каждой комбинации. По О. Моор, 1932
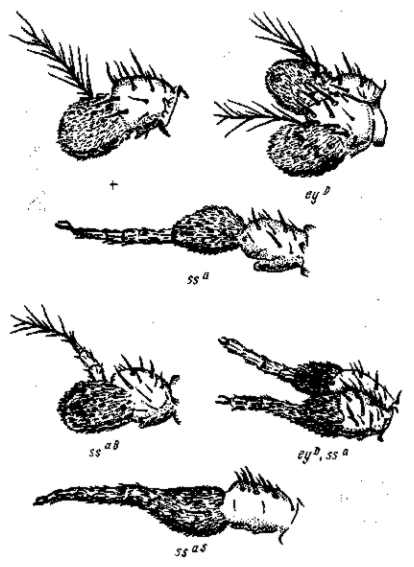
Рис. 16. Строение антенн различных аллеломорфов aristopedia и их комбинаций. Первый рисунок (+) показывает нормальную антенну «дикого» типа Drosophila тelanogaster. По К. Уоддингтону
Если отдельная мутация означает нарушение формообразовательного механизма, то простое суммирование их может вести только к его полному разрушению (рис. 15, 16).
Прежде всего — это само собой разумеется — бесконтрольное суммирование мутаций означает разрушение наследственного аппарата вместе с его защитными механизмами («генный баланс», доминирование нормы и т. и.). Во-вторых, это означает разрушение регуляторного механизма индивидуального развития, защищающего его нормальное течение от возможных нарушений (о чем только что было сказано). И, наконец, это означает разрушение вообще всего реакционного и корреляционного аппаратов, т. е. всей системы факторов индивидуального развития, ведущее к полной дезорганизации, к уничтожению приспособленности организма к внешней среде и к нарушению внутренней согласованности частей и функций. Такой распад организации наблюдается всегда в отношении тех органов и приспособлений, которые в процессе эволюции теряют свое значение. Редукция органов объясняется именно бесконтрольным накоплением мутаций (Шмальгаузен, 1938а). Если этого не происходит в процессе обычного мутирования любого вида организмов, то это объясняется постоянным действием естественного отбора, вводящего этот процесс в определенное русло.
|
|
|
А. СКОРОСТЬ ЕСТЕСТВЕННОГО ОТБОРА
Возможность естественного отбора определяется, конечно, наличием материала для отбора, т.. е. генетическим многообразием особей данной популяции. Максимально возможная скорость естественного отбора определяется масштабом изменчивости в признаках, имеющих какое-либо значение в борьбе за существование. В математическом выражении эта зависимость известна как основной закон селекции Р. Фишера (R. Fisher). По этому закону скорость отбора признака пропорциональна квадрату среднего квадратического уклонения данного признака, т. е. варианте. Изменчивость особей популяции зависит от ее насыщенности различными мутациями, которая бывает различной для разных мутаций в зависимости от частоты их возникновения и от биологического их значения. Большинство мутаций неблагоприятны для их обладателей, но некоторые из них могут давать частичные преимущества в некоторых локальных, сезонных или просто случайных условиях существования. Однако даже безусловно вредные мутации, если они рецессивны, могут широко распространиться в популяции. Во всех случаях распространение мутаций зависит от частоты повторного воьникновения данной мутации (мутабильности) и скорости ее уничтожения под влиянием естественного отбора (при полной рецессивности только после появления гомозиготных мутаций). Между мутационным «давлением» и «давлением» естественного отбора устанавливается некоторое равновесие на разном уровне концентрации для разных мутаций — более высоком для условно благоприятных или «индифферентных» мутаций и более низком для вредных и в особенности для полудоминантных деталей. Такое разнообразие в генетическом составе популяции и, особенно, большое число гетерозигот по разным мутациям составляют как бы основной фонд, или резерв наследственной изменчивости, служащий очень благоприятной основой для эффективного действия естественного отбора и быстрой перестройки генетической структуры популяции. По мере этого некоторые мутации элиминируются, другие, наоборот, суммируются, частью переходят в гомозиготное состояние и входят в состав новой нормы. Скорость естественного отбора зависит не только от изменчивости популяции, т. е. резерва уже накопленных изменений, но и от скорости мутирования. Возникновение неблагоприятных мутаций замедляет течение отбора, а возникновение благоприятных ускоряет его. Однако скорость мутирования вообще очень незначительна, да и отбору подлежат не отдельные мутации, а сложные их. комбинации в виде тех или иных фенотипов. Число таких комбинаций, конечно, неизмеримо больше числа мутаций и о недостатке материала для отбора, идущего всегда по фенотипам, не может быть и речи. Во всяком случае мутирование при определенной его скорости, хотя и может повлиять на общую скорость естественного отбора, не может изменить его направления и закономерного распределения скоростей в зависимости от величины коэффициента селекции и от концентрации отбираемых вариантов. Равномерное мутирование может привести лишь к вычитанию или прибавлению некоторой постоянной величины к приводимым далее значениям скоростей отбора.
|
|
|
Вопрос об эффективности естественного отбора теоретически разработан в целом ряде исследований Р. Фишера, Дж. Холдэна и С. Райта (R. Fisher, J. Haldane, S. Wright). Математический аппарат этих работ столь сложен, что не допускает доступного изложения. Мы отметим лишь немногие выводы. Прежде всего укажем, что по сделанным расчетам даже самые малые селекционные преимущества могут быстро привести к весьма заметным сдвигам в строении популяции. Большое значение имеют размеры популяции. В малых популяциях происходят родственные скрещивания, легко приводящие, при случайностях в элиминации, как к случайной утрате, так и к случайному фиксированию генов в гомозиготном состоянии. Эти случайные явления ускользают из-под контроля естественного отбора и, следовательно, естественный отбор в малых популяциях менее эффективен. Те же явления могут, однако, привести к возникновению новых комбинаций. В больших популяциях имеются гораздо более благоприятные условия для накопления самых разнообразных наследственных изменений. Случайные процессы практически никакой роли не играют, и возможности быстрого отбора очень велики. В больших популяциях, однако, весьма затруднено фиксирование благоприятных комбинаций. Очень большое значение имеют колебания численности популяции. В период максимального сокращения популяции родственные скрещивания ведут к гомозиготизации и фиксированию различных мутаций и их комбинаций в местах переживания остатков популяции. В период размножения естественный отбор ослаблен. Локальные популяции свободно размножаются, скрещиваются и дают начало множеству новых комбинаций, которые затем в период сокращения численности подвергаются самому жесткому отбору. Наиболее благоприятны для быстрого действия естественного отбора именно такие популяции с колеблющейся численностью или малые лишь частично изолированные популяции, которые систематически обмениваются мигрантами (...)
|
|
|
Кривая, выражающая изменение скорости отбора в зависимости от концентрации отбираемого варианта, имеет при малом коэффициенте селекции почти симметричную форму и, следовательно, наибольшая скорость отбора наблюдается на средних его концентрациях. При увеличении коэффициента селекции максимум скорости отбора сдвигается в сторону меньших концентраций и кривая распределения скоростей отбора приобретает асимметричную форму.
У диплоидного организма соотношения становятся более сложными, так как появляется третий, промежуточный тип, который может иметь различное селекционное значение. Соотношение трех типов в равновесной популяции определяется формулой Вейнберга — Гарди: (р + q)2 = р2 + 2pq + q2 = 1. В этом случае при простых мутациях имеются три генотипа АА, Аа и аа, которым соответствуют и фенотипы в числовом соотношении, указанном формулой. При определении скорости отбора, который идет всегда по фенотипам, приходится учитывать не только селекционное преимущество отбираемой гомозиготы, но и гетерозиготы (если она имеет хоть какое-либо отличие от элиминируемого варианта). Это усложняет расчеты, в которых теперь участвуют уже два коэффициента селекции. Ввиду обычного доминирования нормы мы можем рассмотреть лишь этот частный случай, что значительно упростит нашу задачу.
|
|
|
Скорость отбора находится в той же зависимости от селекционного коэффициента и от концентрации отбираемого варианта (фенотипа), как и в случае гаплоидов. Однако сама концентрация вариантов меняется иначе. В данном частном случае, при полном доминировании, гетерозиготы сходны с доминантными гомозиготами и это значительно увеличивает концентрацию доминантного фенотипа. Это ведет к ускорению отбора доминанты, особенно на малых концентрациях гомозиготы. Наоборот, концентрация рецессивного фенотипа оказывается уменьшенной, так как он в генотипе Аа не получает выражения и не может быть предметом отбора.
Таким образом кривые, выражающие распределение скоростей отбора фенотипов в зависимости от концентрации соответствующих генов, получают для доминанты и для рецессива различную форму. В особенности интересен, однако, частный случай отбора при наличии селекционного преимущества гетерозиготы перед обоими гомозиготами. При малом селекционном преимуществе гетерозиготы кривая распределения скоростей отличается от соответствующих кривых для гомозигот лишь большей растянутостью максимума, лежащего на средних концентрациях. При более остром отборе кривая приобретает однако ясно седлообразную форму, т. е. двухвершинный характер. В случае равного преимущества гетерозиготы перед обеими гомозиготами эта кривая оказывается вполне симметричной.
Приведенные формулы, а также кривые распределения скоростей отбора в зависимости от концентрации отбираемого гена в популяции позволяют сделать некоторые интересные заключения. Скорость естественного отбора зависит как от концентрации отбираемого варианта (р), так и от концентрации элиминируемого варианта (д) или, иными словами, она пропорциональна интенсивности соревнования, которую мы измеряем произведением обеих концентраций (w = apq). Скорость отбора зависит также от селекционного преимущества одного варианта перед другим. При малом коэффициенте селекции скорость отбора просто пропорциональна этой величине. При более значительных различиях в конкурентоспособности зависимость сложнее и выражается в нашей формуле величиной s/(1-qs).
Если рассматривать скорость отбора в зависимости от концентрации (р) отбираемого фенотипа, то ясно, что перед нами прямая пропорциональность Δp=p(qs/(1-qs)). В этом случае можно сказать, что скорость отбора определяется не только концентрацией отбираемого варианта, но и его оценкой, которая дается в нашей формуле величиной qs/(1-qs). Оценка отбираемого варианта зависит как от величины коэффициента селекции, так иот концентрации элиминируемого варианта и достигает максимума при его максимальной концентрации. Это собственно понятно, так как оценка в борьбе за существование имеет сравнительный характер. Вариант А может выявить свое преимущество перед В лишь при совместном существовании обоих вариантов в популяции. Если вариант А редок, то его преимущества перед В сказываются с наибольшей ясностью. Если вариант А стал обычным, то он уже теряет свое выигрышное положение в популяции, в которой особи А преобладают. Если же вариант А завоевал всю популяцию, то отдельная особь А уже не имеет никаких преимуществ перед другими особями этой популяции. Другими словами, качество варианта не имеет абсолютного значения. Оно всегда релятивно. Оценка варианта производится всегда на фоне его альтернативы и является максимальной при решительном преобладании в популяции элиминируемого варианта. Чем значительнее коэффициент селекции (его пределом является s = l), тем выше оценка отбираемого варианта уже на самых малых его концентрациях. Поэтому уровень максимальной скорости отбора сдвигается при возрастании коэффициента селекции со средних концентраций при s>0,l все более в сторону меньших концентраций отбираемого варианта. Кривая распределения скоростей отбора в зависимости от концентрации отбираемого гена приобретает при этом все более асимметричную форму. В пределе, при 5=1, скорость селекции определяется только концентрацией элиминируемого варианта и, следовательно, максимальна уже на самых минимальных концентрациях отбираемого варианта (Ар = q).
Седлообразная форма кривой распределения скоростей отбора гетерозиготного фенотипа в зависимости от концентрации одного из генов является добавочной иллюстрацией сказанного. Кривая эта имеет два максимума — один максимум определяется оценкой гетерозиготы А а на фоне полного преобладания в популяции одной гомозиготы (аа) или одного гена (а), а второй максимум определяется оценкой той же гетерозиготы на фоне полного преобладания другой гомозиготы (АА) или гена А. Чем выше значение коэффициента селекции, тем шире раздвинуты оба максимума и тем яснее они выражены.
Все это имеет глубокий биологический смысл. Естественный отбор есть результат внутривидового соревнования особей (вариантов). Если скорость естественного отбора определяется при прочих равных условиях произведением из концентраций обоих вариантов, то это значит, что она пропорциональна напряженности их соревнования. Скорость естественного отбора пропорциональна, следовательно, математической вероятности столкновения обоих альтернативных вариантов. В начале отбора, когда концентрация отбираемого варианта А очень мала, каждая особь А сталкивается практически только с особями В и при этом полностью выявляются ее преимущества. Однако скорость естественного отбора мала, так как число особей А (а следовательно и общее число гетерогенных столкновений) еще очень мало. По мере нарастания концентрации особи А все чаще сталкиваются и с другими особями А и в этом случае не выявляют своих преимуществ. Однако скорость отбора нарастает, так как число особей А, а вместе с тем и абсолютное число столкновений между разными вариантами продолжают нарастать. Когда концентрация А достигает 50%, особи А сталкиваются в своей жизни в равном числе случаев как с особями В, так и с особями А. Только в первом случае выявляется их преимущество. Особи А утратили, следовательно, свое исходное преимущество в популяции по меньшей мере на 50%. Однако абсолютное число столкновений теперь максимально. При преобладании в популяции варианта А он сталкивается уже главным образом с особями А и, следовательно, окончательно лишается своих былых преимуществ в популяции. С падением концентрации варианта В уменьшается теперь и число гетерогенных столкновений и скорость отбора быстро падает.
При отборе гетерозиготы Аа максимум скорости отбора определяется выявлением ее преимуществ при наибольшей вероятности столкновения с гомозиготой аа в случае ее преобладания в популяции и вероятностью столкновения с гомозиготой АА при преобладании последней.
Таким образом естественный отбор определяется в своей скорости внутривидовым соревнованием вариантов, интенсивность которого зависит от распределения концентраций, т. е. от вероятности столкновения альтернативных форм между собой. Кроме этого, скорость естественного отбора определяется, конечно, селекционным преимуществом, выявляемым при сопоставлении этих вариантов в условиях данного биогеоценоза.
Кривая распределения скоростей отбора гетерозиготы замечательна не только по своей форме. Она демонстрирует большую эффективность естественного отбора гетерозиготного фенотипа в огромном диапазоне концентраций каждого из генов (от 0,1 до 0,9) или каждого из гомозиготных фенов (от 0,01 до 0,8).
Мы говорили пока только о скорости отбора фенотипов. Однако отбор фенотипов по необходимости связан и с отбором генотипов.
При отборе гаплоидов отбор фенотипа равнозначен отбору генотипа. В случае диплоидного организма это не совсем так. При полном доминировании скорость отбора доминантного гена снижается вследствие отбора гетерозиготных особей Аа, которые содержат в себе, кроме отбираемого гена А, также элиминируемый ген а. Снижается также скорость отбора рецессивного гена а, который отбирается только по относительно редкому фену аа и оказывается недоступным для отбора по обычным фенам Аа, Для отбора доминантных генов это уменьшение скорости отбора особенно значительно (примерно вдвое) вначале, когда почти все гены А заключаются в гетерозиготах. Однако по мере увеличения количества гомозиготных доминантов отбора гена А быстро возрастает, приближаясь к скорости отбора фена (АА + Аа), так как относительное число гетерозигот в популяции уменьшается. Наоборот, при отборе рецессивного фенотипа аа, отбор гена а идет вначале почти с той же скоростью, как и отбор фена аа, так как последний содержит только гены а. Однако по мере увеличения концентрации а, элиминируемые гены А сохраняются главным образом в гетерозиготах Аа, в которых гены а недоступны для отбора. Элиминация гетерозигот замедляет отбор генов а примерно вдвое (по сравнению со скоростью отбора фенотипа аа).
Для непосредственного определения изменения концентрации генов в популяции (за одно поколение) в результате естественного отбора диплоидных особей по их фенотипам была предложена следующая приближенная формула (Wright): Ар ~ pqs [q -r-+ hs (1– 2q)], где р означает концентрацию доминантного гена А, q — концентрацию рецессивного гена a, s — селекционное преимущество фенотипа АА и hs — селекционное преимущество гетерозиготного фенотипа Аа перед гомозиготой аа. Эта формула пригодна только при малом значении селекционных коэффициентов s и hs и не малых концентрациях р (А).
При равенстве селекционного преимущества гетерозиготы и нормальной гомозиготы АА и, в частности, в случае полного доминирования нормы, вполне точные результаты дает формула Δp=pq2(s/1-q2s)), которая при малых значениях sдает (как и первая формула) приближенно: Δp≈pq2s.
Во всех наших рассуждениях мы считали, что коэффициент селекции является величиной постоянной для данной пары альтернативных вариантов. Это вряд ли так. При отборе положительного варианта А не только нарастает его концентрация, но при постоянном перекомбинировании могут улучшаться и его качества, в результате чего значение коэффициента селекции будет нарастать и это может привести к заметному ускорению отбора. Вероятно, такое ускорение будет наибольшим именно в первой фазе завоевания своего места в системе данной популяции.
Мы видели, что скорость отбора доминантного фена (и гена) возрастает при увеличении концентрации соответствующего гена в популяции вначале очень быстро. Наоборот, скорость естественного отбора рецессивного фена (и гена) нарастает вначале, при малых концентрациях, очень медленно, хотя и достигает позднее того же уровня, как и в случае доминантности. Это ведет при отборе в течение ряда поколений к очень большому отставанию в достижении заметного эффекта при отборе по рецессивным признакам. Однако конечный результат оказывается через некоторое время тем же самым.
Как бы ни было велико значение подобных теоретических расчетов, следует всегда помнить, что их правильность зависит от правильности принятых предпосылок и их значение поэтому всегда условно. Всегда в основу расчетов кладутся результаты известной схематизации, упрощения реальных соотношений. В действительности, например, не существует ни абсолютного доминирования, ни полной рецессивности. Следовательно, реальные кривые скорости отбора займут промежуточное положение между нашими теоретическими кривыми. Совершенно новые мутации не бывают рецессивными и если их выражение хотя бы в какой-либо мере благоприятно, то одновременно с их отбором начнется и отбор на усиление этого выражения в гетерозиготе. Мутация начнет доминировать и это быстро приведет к ускорению отбора, не только вследствие самой доминантности, но и вследствие возрастания коэффициента селекции. Кроме того, в природных условиях жизни амфимиктических популяций эволюция редко строится непосредственно на отборе вновь возникающих отдельных благоприятных мутаций. Обычно популяции находятся в состоянии некоторого генетического равновесия и при изменении условий существования переходят из одного стационарного состояния в другое за счет мобилизации внутренних резервов изменчивости. В этом случае все преимущества будут на стороне рецессивных генов, которые уже обладают, в отличие от доминантных, значительной концентрацией в любой природной популяции. При эволюционной перестройке популяции доминантные и рецессивные мутации стартуют на совершенно разных уровнях концентрации. Поэтому отбор рецессивных мутаций может произойти со значительно большей скоростью, чем отбор доминантных.
В случае выявления каких-либо преимуществ у гетерозиготы ее отбор произойдет именно в начале с исключительно большой скоростью, так как все мутации находятся в генетическом резерве равновесной популяции именно в гетерозиготном состоянии и нередко достигают высокой концентрации. Однако при свободных скрещиваниях и отсутствии специальных генетических механизмов, поддерживающих постоянную гетерозиготность, полное насыщение популяции гетерозиготной мутацией оказывается недостижимым (максимальный уровень насыщения достигается только при малой жизнеспособности гомозиготной мутации).
Б. КОМБИНИРОВАНИЕ МУТАЦИЙ
При половом размножении происходит не только распространение мутаций в потомстве, а следовательно — и в популяции, но и их разнообразное комбинирование. При свободном скрещивании (панмиксии) особей, обладающих генетическими различиями, создаются новые комбинации, которые в потомстве расщепляются и затем опять комбинируются, создавая при каждом скрещивании все новые и новые комбинации (рис. 15, 16). Если бы все комбинации были жизнеспособны и равноценны, то их возможное число достигло бы величин, поражающих воображение. Если бы генотип организма содержал лишь 1000 разных генов (несомненно, что у дрозофилы их больше) и каждый ген был способен к десяти изменениям (т. е. мог бы дать начало десяти аллеломорфам), то число возможных комбинаций было бы равно 101000, т. е. выражалось бы сверхастрономическим числом. Эти числа, конечно, не могут нигде реализоваться. Огромное большинство комбинаций было бы абсолютно нежизненным. Ничтожная доля возможных комбинаций все еще невероятно разнообразна и может быть представлена всеми переходами от полной летальности до высшего уровня приспособленности, достигнутой данным организмом. Можно эти реально возможные комбинации представить себе, по Райту, распределенными по полю, на котором низменности и «долины» означают низшие уровни жизнеспособности, а возвышенности с их «вершинами» соответствуют высшим уровням адаптации организма. В больших нерасчлененных популяциях имеется одна такая главная возвышенность, которая в случае падающей мутабильности и возрастающей интенсивности отбора сокращает свою площадь и становится более крутой, поднимая свою вершину до максимального уровня приспособленности. Такое состояние соответствует специализации.
При этих условиях возможность использования новых мутаций, которые лишь редко попадают в область главной возвышенности, очень низка, и такие виды могут при изменении условий оказаться недостаточно пластичными. Изменение факторов среды ведет к изменению «рельефа» площади, занимаемой комбинациями генов, и к постепенному перемещению вершины приспособленности в известном направлении, соответственно новым условиям существования.
Сокращение величины популяции ведет к уменьшению числа возможных комбинаций, к уменьшению изменчивости, к «фиксации генов» более или менее независимо от их адаптивной ценности. Поле комбинаций сокращается; «вершина» уменьшается и блуждает без определенного направления. Это — путь вымирания. Наконец, при разбивке вида на различные формы и местные расы, которые обладают своими более или менее обособленными вершинами приспособленности, судьба отдельных возвышенностей может быть весьма различной: одни из них сокращаются и исчезают, другие возникают вновь на месте прежних «долин», третьи перемещаются и расширяются или достигают высшего уровня специализации. Однако для вида в целом создаются, благодаря комбинации внутри группового и межгруппового отборов, наиболее благоприятные условия для прогрессивной эволюции, в особенности если отдельные популяции не вполне изолированы и постоянно обмениваются мигрантами.
Известный баланс между факторами эволюции («не слишком высокая мутабильность, не слишком жесткий отбор, не слишком тесный инбридинг и не слишком широкое скрещивание») является основным условием пластичности организмов (С. Райт).
В. ОГРАНИЧЕНИЕ ПАНМИКСИИ
Изоляция (постоянное ограничение панмиксии). Территориальное или биологическое ограничение скрещивания, раз установившись, сохраняет свое значение длительно и обычно со временем все более углубляется. Начинаясь как частичное ограничение свободы скрещивания, нарушаемое постоянной миграцией отдельных особей, оно дополняется, по мере установления различий между обособленными популяциями, постепенным развитием более существенных преград, рано или поздно полностью устраняющих возможность скрещивания. Наиболее общее значение имеет бесспорно дарвиновский фактор — элиминация менее приспособленных особей, устраняемых тем самым от возможности скрещивания с преуспевающими особями, которые в данных условиях оказались более (приспособленными. Из других возможных форм изоляции наиболее общий характер имеет, по-видимому, территориально-географическое расчленение, особенно существенное на начальных этапах обособления новых форм. На дальнейших этапах расхождения оно переходит в различные формы биологической изоляции или дополняется ими. Из этих последних следует учитывать значение экологической изоляции, не всегда отделимой от географической, значение различных форм физиологической (в том числе — половой) изоляции и, наконец, значение генетической изоляции, которая, проявляясь в стерильности гибридов, иногда устанавливается сразу, но чаще всего завершает процесс полного обособления новой формы от исходной (т. е. процесс видообразования).
Мы здесь рассмотрим значение раличных форм изоляции лишь как факторов, способствующих начальному сохранению и распространению новых комбинаций через ограничение их скрещивания с исходными формами, оставив в стороне большой вопрос о значении изоляции для видообразования и для дальнейшего расхождения органических форм.
1. Территориальная изоляция. Каково бы ни было значение географических преград, в виде непроходимых для известного вида пространств суши или воды, горных хребтов, пустынь, рек и т. п., мы не имеем в виду разбирать здесь эти вопросы, как общеизвестные и имеющие вместе с тем лишь частное значение в видообразовании. Мы остановимся на менее заметном и менее полном территориальном разграничении, которое имеет, однако, более общее значение, в особенности в начальных процессах дифференциации отдельных популяций в пределах вида или подвида. Вместе с тем как частичное, допускающее довольно широкий обмен особями между отдельными популяциями, оно имеет в большей мере прогрессивное значение, способствуя сохранению пластичности и возможности более быстрых темпов эволюции.
Такое частичное ограничение скрещивания достигается уже наличием самого факта размещения отдельных особей в пространстве, в особенности у малоподвижных или оседлых (хотя бы лишь в период размножения) животных. Его значение наиболее ясно у сидячих животных и у растений.
Сошлемся прежде всего на образование целых рядов форм в тех случаях, когда известный род или вид занимает большой ареал распространения и лишен возможности свободного перемещения в пределах этого ареала. В особенности это касается многих растений. Хорошим примером может служить сон-прострел (Anemone Pulsatilla), который широко распространен по континенту Евразии и дает непрерывный ряд форм, закономерно изменяющихся в направлении с запада на восток Европы. На западе подвид Л. p. ssp. germanica обладает тонко рассеченными и разбросанными листьями и поникшими цветами. На востоке у подвида А. р. ssp. grandis более грубо рассеченные, стоячие листья и стоячие цветы. Этот комплекс изменений связан с обилием дождей на западе и засушливостью восточной Европы (Zimmermann, 1938). Положение листьев способствует стеканию дождевой воды к корневой системе у восточных форм и, наоборот,— разбрызгиванию воды — у западных. Поникшее положение цветов предохраняет их от смывания пыльцы дождевой водой. Степень рассеченности листьев связана с различной интенсивностью транспирации. Мало рассеченные листья восточной формы испаряют заметно меньше воды, чем сильно рассеченные листья западной формы (что доказано измерением транспирации в одинаковых условиях опыта). Приспособительность различий совершенно ясна, но вместе с тем — местные расы образуют еще совершенно непрерывный ряд форм в пределах вида между обоими крайними подвидами. Различия оказываются наследственными и типично расщепляются в потомстве гибридов. Однако одновременно имеются и модифика-ционные изменения, которые наслаиваются на наследственные.
Рыба бельдюга (Zoarces viviparus) образует в Скандинавии, по Шмидту, в отдельных фиордах совершенно непрывные ряды форм. В направлении от устья к концу фиорда у этой рыбы постепенно уменьшается число позвонков и число лучей некоторых плавников.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 526; Нарушение авторских прав?; Мы поможем в написании вашей работы!