КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Экология сообществ и экосистем 4 страница
С понятием экологической ниши тесно связано представление о так называемых насыщенных и ненасыщенных биоценозах. Первые представляют собой экосистемы, в которых жизненные ресурсы на каждом этапе преобразования биомассы и энергии используются наиболее полно. Когда жизненные ресурсы утилизируются частично, биоценозы можно называть ненасыщенными. Для них характерно наличие свободных экологических ниш. Однако это в высшей степени условно, поскольку экологические ниши не могут существовать сами по себе, независимо от занимающих их видов. В противном случае неточным было бы определение биоценоза, где он выступает как система, состоящая из экологических ниш, занятых теми или иными организмами.
Ненасыщенные биоценозы обладают потенциальной способностью принять в свой состав новые виды, которые, заняв определенные экологические ниши, будут содействовать наиболее полной утилизации жизненных ресурсов сообщества. Но так как неиспользованные резервы, нереализованные возможности интенсификации потока веществ и энергии имеются практически в любом биогеоценозе (иначе не могло бы осуществляться их постоянное развитие во времени и пространстве!), все биоценозы условно можно считать ненасыщенными. Чем меньше насыщенность биоценоза, тем легче вводятся в его состав и удачнее акклиматизируются новые виды.
Видовая структура. Каждый конкретный биоценоз характеризуется строго определенным видовым составом. При этом одни виды биоценоза могут быть представлены многочисленными популяциями, а другие малочисленными. В связи с этим в любом биоценозе можно выделить один или несколько видов, определяющих, его облик.
Типичной характеристикой биоценоза является относительна малое количество видов, особи которых встречаются часто (представлены большим числом особей). И наоборот, в каждом биоценозе есть много видов с небольшим числом особей. Так, облик лесного либо степного биоценоза представлен одним или несколькими видами растений. В дубраве – это дуб, в бору – сосна, в ковыльно-типчаковой степи ·– ковыль и типчак. В лесу, состоящем из десятков видов растений, только один или два из них дают до 90 % древесины. Эти виды называются доминирующим и или доминантными. Они занимают ведущее, господствующее, положение в биоценозе. Обычно наземные биоценозы называют по доминирующим видам: лиственничный лес, сфагновое болото, ковыльно-типчаковая степь.
Виды, живущие за счет доминантов, получили название предоминантов. К примеру, в дубовом лесу таковыми являются кормящиеся на дубе насекомые, сойки, мышевидные грызуны.
В биоценозе есть и так называемые эдификаторы. Это строители сообщества, т. е. виды, создающие условия для жизни других видов данного биоценоза. Рассмотрим эдификаторную роль ели и сосны.
Ель в таежной зоне образует густые, сильно затененные леса. Под пологом ее могут обитать только растения, приспособленные к условиям сильного затенения, повышенной влажности воздуха, кислых оподзоленных почв. Соответственно этим факторам в еловых лесах формируется и специфичное животное население. Следовательно, ель в данном случае выступает в роли мощного эдификатора, обусловливающего определенный биоценоз.
В сосновых лесах эдификатором является сосна. Но по сравнению с елью она – более слабый эдификатор, поскольку сосновый лес относительно светлый и редко-ствольный. Его видовой состав растений и животных гораздо богаче и разнообразнее, чем в ельнике. В борах встречаются даже такие растения, которые могут жить вне леса.
Виды-эдификаторы встречаются практически в любом биоценозе. На сфагновых торфяниках это сфагновые мхи. Они создают специфичные условия биоценоза, которые отличаются плохой аэрацией и низкой теплопроводностью торфа, кислой реакцией среды, бедностью элементов минерального питания для высших растений. В степных биоценозах мощным эдификатором служит ковыль. Однако роль эдификаторов в тех или иных биоценозах не абсолютна. Она зависит от многих факторов. Так, при изреживании елового леса ель может утратить функции мощного эдификатора, поскольку при этом происходит осветление леса и в него внедряются другие породы, снижающие эдификаторное значение ели. В сосняке на сфагновых болотах сосна также теряет свое эдификаторное значение. Его приобретают сфагновые мхи.
Все виды, слагающие биоценоз, в определенной степени связаны с доминирующими видами и эдификаторами. Внутри биоценоза формируются более или менее тесные группировки, комплексы популяций, зависящие либо от растений-эдификаторов, либо от других элементов биоценоза. Так создаются своеобразные структурные единицы биоценоза – консорции. Наиболее детально учение о консорциях разработали В. Н. Беклемишев и Л. Г. Раменский.
Консорция – это совокупность популяций организмов, жизнедеятельность которых в пределах одного биогеоценоза трофически или топически связана с центральным видом – автотрофным растением. Обычно в роли центрального вида выступает эдификатор – основной вид, определяющий особенности биоценоза. Популяции остальных видов консорции образуют ее ядро, за счет которого существуют виды, разрушающие органическое вещество, создаваемое автотрофами. Примером консорции может служить растение со всеми связанными с ним организмами (эпифиты, паразиты, вредители, симбионты).
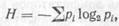
|
Однако, как известно, видовая структура биоценоза характеризуется не только числом видов, входящих в его состав, т. е. видовым разнообразием, но и их количественным соотношением. Например, если в двух сравниваемых биоценозах растения представлены двумя видами и в каком-то из них 90 % особей принадлежит одному виду, тогда как в другом на долю особей каждого вида приходится по 50 %, то эти биоценозы будут сильно отличаться друг от друга. Количественное соотношение видов в биоценозе называется индексом разнообразия (Я) и чаще всего определяется по формуле Шеннона:
где pi – доля каждого вида в сообществе.
Пространственная структура биоценоза. Первоначальное заселение организмами того или иного биотопа определяется его экологическими факторами и прежде всего особенностями атмосферы, горной породы, почвы и ее вод. Ярусность расположения их накладывает отпечаток и на распределение организмов в биотопе. В ходе длительного эволюционного преобразования, приспосабливаясь к определенным абиотическим и биотическим условиям, живые организмы в итоге так разместились в биогеоценозе, что практически не мешают друг другу, т. е. их распределение носит ярусный характер.
Ярусность – это явление вертикального расслоения биоценозов на разновысокие структурные части. Наиболее четко она выражена в растительных сообществах (фитоценозах). Благодаря ярусности различные растения, особенно их органы питания (листья, окончания корней), располагаются на разной высоте (или глубине) и поэтому легко уживаются в сообществе. Это способствует увеличению числа организмов на единицу площади, значительному ослаблению конкуренции между ними, более полному и разностороннему использованию условий среды.
Фитоценоз приобретает ярусный характер, при наличии в нем растений, различающихся по высоте.
В лесу, например, нередко выделяется до 6 ярусов: I – деревья первой величины (ель, сосна, дуб, береза, осина); II–деревья второй величины (рябина, черемуха); III – подлесок из кустарников (лещина, бересклет, шиповник); IV – подлесок из высоких кустарничков и крупных трав (багульник, голубика, вереск, аконит, иван-чай); V–низкие кустарнички и мелкие травы (водяника, клюква, кисличка); VI – мхи, напочвенные лишайники, печеночники.
Ярусно располагаются и подземные части растений. Корни у деревьев, как правило, проникают на большую глубину, чем у кустарников, ближе к поверхности располагаются корни мелких травянистых растении, а непосредственно на ней – ризоиды мхов. При этом в поверхностных слоях почвы корней значительно больше, чем в глубинных (см. рис. 20).
Ярусы определяют сложение и структуру фитоценоза. Если их мало, то растительное сообщество называют простым; много – сложным.
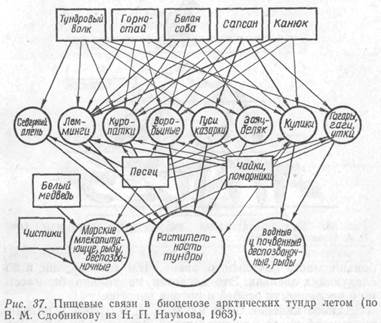
Растения каждого яруса и обусловленный ими микроклимат создают определенную среду для специфичных животных. В конечном итоге возникают группировки растений и животных – популяции тесно связанных между собой организмов. В почвенном ярусе леса, заполненном корнями растений, обитают бактерии, грибы, насекомые, клещи, черви. В лесной подстилке среди разлагающихся растительных остатков, мхов, лишайников и грибов живут насекомые, клещи, пауки, множество микроорганизмов. Более высокие ярусы – травостой, подлесок – занимают растительноядные насекомые, птицы, млекопитающие и другие животные. При этом даже птицы, свободно передвигающиеся, обычно придерживаются строго определенного яруса. Особенно ярко это проявляется в гнездовой период (рис. 36).
 Следовательно, ярусы в биоценозе различаются не только высотой, но и составом организмов, их экологией и той ролью, которую они играют в жизни всего сообщества. Одни и те же виды в одном сообществе в силу возрастных различий особей или частичного угнетения могут находиться весьма кратковременно в разных ярусах. Например, всходы ели, пока они маленькие, располагаются в нижних ярусах леса. Но по мере роста при благоприятных условиях ель займет свое место в верхнем ярусе. Однако зная экологию ели, ее даже в молодом возрасте нельзя относить к растениям нижних ярусов.
Следовательно, ярусы в биоценозе различаются не только высотой, но и составом организмов, их экологией и той ролью, которую они играют в жизни всего сообщества. Одни и те же виды в одном сообществе в силу возрастных различий особей или частичного угнетения могут находиться весьма кратковременно в разных ярусах. Например, всходы ели, пока они маленькие, располагаются в нижних ярусах леса. Но по мере роста при благоприятных условиях ель займет свое место в верхнем ярусе. Однако зная экологию ели, ее даже в молодом возрасте нельзя относить к растениям нижних ярусов.
Следует отметить, что имеются и внеярусные организмы. Это лианы, различные эпифиты, паразиты, а также многие животные, свободно переходящие из одного яруса в другой. Они затрудняют четкое выделение ярусов, что особенно выражено в тропических влажных лесах, структура которых чрезвычайно сложна.
Виды животных и растений разных ярусов в большинстве случаев существенно различаются по своему отношению к условиям среды. Закономерно, что растения каждого нижележащего яруса более теневыносливы, чем расположенные над ними. Связано это с тем, что освещение при переходе от верхних ярусов к нижним все более ослабевает и для световых растений становится недостаточным. При этом виды различных ярусов в биоценозе находятся в тесном взаимоотношении и взаимозависимости. Сильное разрастание верхних ярусов сообщества соответственно уменьшает густоту нижних, нередко вплоть до полного исчезновения слагающих их растений. Вместе с ними исчезает и животное население.
С другой стороны, разреживание верхнего яруса по тем или иным причинам способствует усиленному развитию растений нижних ярусов, благодаря улучшению режима света, влаги, тепла, а также повышению содержания минеральных веществ в почве. Разрастание нижних ярусов положительно влияет на животное население как в количественном, так и в качественном отношении. Особенно ярко это выражено в лесных биоценозах, где в случае разреживания древостоя обильно разрастаются кустарники или светолюбивая травянистая растительность, а при полном слиянии древостоя, к примеру в молодняках, иногда подавляются даже самые теневыносливые травы и мхи. Однако не всегда верхний ярус доминирует над нижним. На верховом болоте, например, господствующая роль принадлежит сфагновому покрову.
Следовательно, ярус можно рассматривать как структурную единицу биоценоза, отличающуюся от других частей его определенными экологическими условиями и набором растений, животных и микроорганизмов. В каждом ярусе складывается своя система взаимоотношений составляющих компонентов.
Вертикальное распределение организмов в биоценозе обусловливает и определенную структуру в горизонтальном направлении. Такие структурные части биоценоза получили название синузий. Они образуются потому, что растения, распределяясь неравномерно, создают то большие, то меньшие скопления (сгущения) и придают тем самым растительному покрову своеобразный мозаичный характер. Между ярусами (если они четко выражены и в них ясно прослеживается специфичная система взаимоотношений слагающих его компонентов) и синузиями много общего.
В геоботанике синузия рассматривается как структурная часть фитоценоза, характеризующаяся определенным видовым составом и эколого-биологическим единством входящих в нее видов. Это, например, синузия сосны, синузия брусники, синузия зеленых мхов и другие синузий лесной зоны. В полынно-солянковой пустыне можно выделить синузию ранневесенних эфемеров или эфемероидов, синузий летне-осенних кустарничков (полыни, солянки).
В лесу неравномерность древесного полога особенно сильно отражается на нижележащих ярусах – на их животном населении, почве, лесной подстилке, микробном составе, а также на климате. В этом случае синузии называются парцеллами. Π а р ц е л лы – это структурные части горизонтального расчленения биогеоценоза, отличающиеся составом, структурой, свойствами компонентов, спецификой их связей и материально-энергетического обмена. В отличие от синузии и яруса, преимущественно геоботанических понятий, парцелла – комплексная единица, так как на правах участников обмена веществ и энергии в нее входят растения, животные, микроорганизмы, почва, атмосфера.
Экологическая структура. Каждый биоценоз слагается из определенных экологических групп организмов. Эти группы, занимая сходные экологические ниши, в разных биоценозах могут иметь неодинаковый видовой состав. Например, в сухих аридных условиях доминируют склерофиты и суккуленты, а на увлажненных территориях – гигрофиты. Экологическую структуру биоценоза отражает также соотношение групп организмов, объединяемых сходным типом питания. Так, в лесах преобладают сапрофаги, в степных и полупустынных зонах ·–· фитофаги, в глубинах Мирового океана – хищники и детритоеды и т. д.
Наиболее ярко различия в экологической структуре биоценоза проявляются при сравнении сообществ организмов в сходных биотопах разных регионов. К примеру, антилопы в саваннах Африки, бизоны в прериях Америки, кенгуру в саваннах Австралии, куница в европейской и соболь в азиатской тайге занимают сходные экологические ниши и выполняют одни и те же функции. Такие виды определяют экологическую структуру сообщества и называются замещающими или викарирующими.
Таким образом, экологическая структура биоценоза – это его состав из экологических групп организмов, выполняющих в сообществе в каждой экологической нише определенные функции.
Экологическая структура биоценоза в комплексе с видовой и пространственной, а также с особенностями экологической ниши служит макроскопической характеристикой его. В отличие от микроскопической характеристики, дающей представление о связях каждой популяции и вида в сообществе вплоть до мельчайшей детали их экологии, макроскопическая характеристика дает возможность определить свойства того или иного биоценоза, выяснить его устойчивость во времени и пространстве, а также предвидеть последствия изменений, вызванных влиянием антропических факторов.
Простые и сложные биоценозы. Биоценоз, как правило, состоит из многих видов. Количество их обычно значительно превышает число видов-доминантов, несмотря на то что последние все же оказывают решающее влияние на характер биоценоза в целом.
Биоценозы различаются видовым разнообразием слагающих их организмов. Часто эти различия очень существенные. Так, в биоценозы тропических лесов входят десятки тысяч видов растений, сотни тысяч видов беспозвоночных и несколько тысяч видов позвоночных животных. Популяции всех видов, составляющих эти биоценозы, очень сложно связаны между собой, а также с физико-химическим окружением. Биоценозы же тундры, пустыни включают несравненно меньшее количество видов. По данным Б. А. Тихомирова, на Таймыре в тундровом биоценозе насчитывается всего 139 видов высших растений, 670 видов низших растений, около 1000 видов животных и 2500 видов микроорганизмов. Естественно, и биомасса, и продуктивность этих биоценозов также различаются, однако взаимосвязи видов, их популяций не столь сложны и многообразны.
На основании взаимоотношений видов в популяциях биоценозы подразделяются на сложные и простые.
Многоярусные биоценозы, состоящие из большого количества популяций многих видов растений, животных и микроорганизмов, связанных между собой разнообразными пищевыми и пространственными отношениями, называются сложными. Сложные биоценозы наиболее устойчивы к неблагоприятным воздействиям. Исчезновение отдельных элементов, например, вымирание какого-либо вида, существенно не отражается на судьбе таких биоценозов, поскольку при этом происходит лишь незначительная перестройка их организации. В исключительно сложных биоценозах тропических лесов никогда не наблюдаются вспышки массового размножения отдельных видов.
Для простых тундровых биоценозов характерны сильные подъемы численности леммингов, падения или взлеты численности песцов и животных, оказывающих существенное влияние па растительный покров. Объясняется это тем, что в упрощенном тундровом биоценозе не хватает видов, которые при необходимости могли бы заменить основной вид и выступить в качестве корма для хищников.
Пограничный эффект. Одним из важнейших признаков структурной характеристики биоценозов является наличие границ сообществ. Однако они очень редко бывают четкими. Обычно соседние биоценозы постепенно переходят один в другой. В результате образуется довольно обширная пограничная зона, отличающаяся особыми условиями. Например, границы между лесом и лугом, лесом и болотом, между лесами с различными видами-эдификаторами выражены хорошо. Однако, когда озеро окружено болотом, переходящим в сухой лес, границы между озером и болотом, болотом и лесом передвигаются в связи с влажностью, которая обычно в течение сезона меняется. И даже независимо от сезонного изменения условий границы между этими биогеоценозами не будут резкими, поскольку растения и животные, характерные для каждого из них, проникают на соседние территории, создавая специфическую «опушку», пограничную полосу, называемую экотоном.
Пограничная зона между двумя биоценозами занимает промежуточное между этими сообществами положение, так как отличается от них температурным режимом, влажностью, освещенностью. В ней как бы переплетаются типичные условия соседствующих биоценозов. Иными словами, в переходной полосе произрастают растения, характерные для обоих биоценозов. Обилие их привлекает сюда и разнообразных животных, так что пограничная зона, как правило, более богата жизнью, чем каждое из смежных сообществ.
Для экотонов характерны так называемые краевые воздействия примыкающих друг к другу биогеоценозов. Они возникают при взаимопроникновении сообществ. Данное явление получило название опушечного эффекта. На опушках наблюдается более быстрая смена растительности, чем в стабильном биоценозе. Так, на вырубках происходит порослевое восстановление леса, а часто и полная перестройка растительного сообщества. Освободившиеся экологические ниши захватываются активно прорастающими растениями, и в это время обостряется конкуренция между отдельными видами. В результате интенсивность животной и растительной жизни на опушках настолько возрастает, что колебания численности популяций выражены здесь гораздо резче, чем в девственном лесу. На опушках выше и плотность популяций по сравнению с соседними стациями.
Однако проникновение того или иного вида в новые биоценозы не всегда сопровождается глубокой перестройкой его организации и выработкой соответствующих адаптации. Как показал немецкий лимнолог А. Тинеманн, определенные структуры и функции некоторых животных дают им возможность заселять новые биотопы, не претерпевая изменений, т. е. эти структуры и функции оказываются адекватными изменившимся условиям. Представления о таких преадаптациях, т. е. принцип «использования преимуществ», вошли в экологическую литературу под названием «правило Тинеманна». Примером реализации этого правила может служить группа брюхоногих моллюсков, у которых слизистая клейкая нога дает им преимущества в смысле возможности широко расселяться как в водоемах, так и в прибрежной полосе.
Различия водных и наземных биоценозов. Структура и функции сообществ рассматривались главным образом на примерах наземных биоценозов. Однако между наземными и водными биоценозами, несмотря на сходное проявление основных закономерностей, имеются и различия, обусловленные разным характером окружающей среды. Структура водных биогеоценозов проще наземных. Это объясняется тем, что во многих водных биогеоценозах, и прежде всего в глубоководных, нет атмосферы и почвы. Несколько сложнее структура мелководных биогеоценозов, так как в прибрежных водных сообществах значительную роль играет и воздушная среда. Донные грунты отличаются от почвы, и связь растений и донных животных с грунтом водоема носит преимущественно механический характер.
На организмы водных биоценозов воздействуют такие факторы среды, которые не имеют никакого значения для организмов наземных сообществ и наоборот. При изучении наземных биогеоценозов прежде всего приходится исследовать влажность и температуру среды, а водных – температуру, газовый режим, растворенные и взвешенные в среде вещества (табл. 8).
В водных биоценозах преобладают низшие растения и в первую очередь водоросли, тогда как в наземных высшие цветковые растения. Животное население водных биоценозов представлено чрезвычайно широко, но высшие членистоногие и высшие позвоночные имеют здесь второстепенное значение. В наземных же биоценозах, наоборот, насекомые (высшие членистоногие) и позвоночные, особенно птицы и млекопитающие, играют главную роль.
Таблица 8. Условия обитания организмов воздушной и водной среды (по Д. Ф. Мордухай-Болтовскому, 1974)
| Условия обитания | Значение условий для организмов | |
| воздушной среды | водной среды | |
| Влажность | Очень важное (часто в дефиците) | Не имеет (всегда в избытке) |
| Плотность среды | Незначительное (за исключением почвы) | Большое по сравнению с ее ролью для обитателей воздушной среды |
| Давление | Почти не имеет | Большое (может достигать 1000 атмосфер) |
| Температура | Существенное (колеблется в очень больших пределах (от –80° до + 100°С и более) | Меньшее по сравнению со значением для обитателей воздушной среды (колеблется гораздо меньше – обычно от –2° до + 40 °С) |
| Кислород | Несущественное (большей частью в избытке) | Существенное (часто в дефиците) |
| Взвешенные вещества | Не важное; не используются в пищу (главным образом минеральные) | Важное (источник пищи; особенно органические вещества) |
| Растворенные вещества в окружающей среде | В некоторой степени (имеют значение только в почвенных растворах) | Важное (в определенном количестве необходимы) |
В водной среде иначе складываются и трофические связи. Здесь важнейшим источником пищи организмов служат взвешенные в толще воды вещества и обитатели планктона, а также детрит, т. е. неполностью разложившиеся продукты распада организмов, в наземных же сообществах преобладают фитофагия и зоофагия.
Между типично водными и полночленными наземными биогеоценозами имеется ряд переходных форм. Например, лишенные почвы болотные биогеоценозы. Более простой компонентной структурой, чем обычные наземные биогеоценозы, обладают биогеоценозы птичьих базаров, так как в них нет почвы и растительности, вследствие чего отсутствует сбалансированный обмен органических веществ. Очень близки по компонентной структуре к наземным биогеоценозам болот и птичьих базаров типичные водные биогеоценозы зарастающих отмелей. Промежуточное положение между водными и наземными биогеоценозами занимают заливные участки пойменных террас и участки морских побережий, подвергающихся воздействию приливов и отливов.
Глава 12. ЭНЕРГЕТИКА ЭКОСИСТЕМ
Цепи и циклы питания. Как уже отмечалось, между организмами биоценоза устанавливаются прочные пищевые взаимоотношения. В результате возникают цепи питания. Они и объединяют прямо или косвенно большую группу организмов в единый комплекс. Цепь питания обычно состоит из трех основных звеньев.
Первое звено образуют так называемые продуценты или производители. Это автотрофные зеленые растения, которые в процессе фотосинтеза создают органическое вещество – первичную биологическую продукцию – и аккумулируют солнечную энергию.
Второе звено представлено консументами, т. е. потребителями,– гетеротрофными организмами, питающимися растениями или другими гетеротрофами. Различают консументы первого порядка (фитофаги), второго порядка (плотоядные животные, питающиеся фитофагами), третьего порядка (хищники, питающиеся другими животными) и т. д.
Третье звено – это редуценты, или деструкторы,– разрушители органического вещества. К ним относятся микроорганизмы, грибы и организмы, питающиеся мертвым органическим веществом и минерализующие его до простых неорганических соединений.
В каждой цепи питания формируются определенные трофические уровни, характеризующиеся различной интенсивностью протекания потока веществ и энергии. Зеленые растения – созидатели органического вещества – образуют первый трофический уровень, фитофаги – второй, плотоядные животные – третий и т. д.
Все звенья цепи питания взаимосвязаны и взаимозависимы. Между ними от первого к последнему осуществляется передача вещества и энергии. Суть этого явления будет рассмотрена ниже. Сейчас важно обратить внимание на то, что при передаче энергии с одного трофического уровня на другой происходит ее потеря. В результате цепь питания не может быть длинной, как это иногда изображают графически. Скорее всего она состоит из 4–6 звеньев (схема 4).
Однако такие цепи в чистом виде в природе обычно не встречаются, поскольку одни и те же виды могут быть одновременно в разных звеньях. Это происходит потому, что монофагов в природе чрезвычайно мало, чаще встречаются олигофаги и полифаги. Рассмотрим, к примеру, цепь питания, основным звеном которой является капуста. Следующим звеном в ней будут гусеницы капустной белянки, капустной моли, капустной совки, зайцы, т. е. все животные, питающиеся капустой. Следовательно, капуста здесь выступает основным звеном многих цепей, поскольку от последующего звена (гусеницы, зайцы и др.) могут тянуться еще цепи. Причем каждый организм, питающийся капустой, одновременно может быть составной частью не одной, а нескольких цепей. Так, заяц, поедая разные растения, входит как консумент первого порядка в большое количество цепей питания. Хищники также питаются различными растительноядными и плотоядными животными, а потому являются звеньями многих цепей.
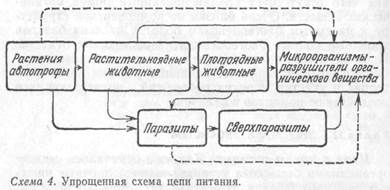
Подобные общие звенья связывают цепи питания в сложную систему. В результате в каждом биоценозе исторически формируются комплексы цепей питания, представляющие собой единое целое. Так создаются циклы, или сети, питания (рис. 37). Если принять во внимание, что практически каждый организм цепи питания выступает в роли хозяина по крайней мере одного, а чаще нескольких паразитов, составляющих в свою очередь звенья других цепей, то нетрудно вообразить всю сложность циклов питания биоценоза.
Экологическая пирамида. В любой цепи питания не вся пища используется на рост особи, т. е. на накопление биомассы. Часть ее расходуется на удовлетворение энергетических затрат организма: на дыхание, движение, размножение, поддержание температуры тела. При этом биомасса одного звена не может быть переработана последующим полностью. В противном случае исчезли бы ресурсы для развития живой материи. В каждом последующем звене пищевой цепи происходит уменьшение биомассы по сравнению с предыдущим. Обычно чем больше масса начального звена, тем она больше в последующих звеньях. Это касается не только биомассы, но и численности особей и потока энергии.
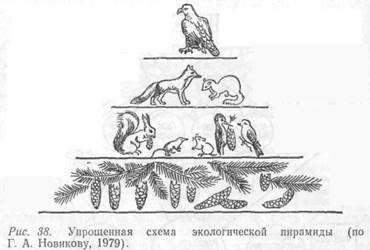
Данное явление было изучено Ч. Элтоном и названо пирамидой чисел или пирамидой Элтона (рис. 38). Различают пирамиду численности (особей), пирамиду биомассы и пирамиду энергии.
Основание пирамиды образуют растения-продуценты. Над ними располагаются фитофаги. Следующее звено представлено консументами второго порядка. И так далее до вершины пирамиды, которую составляют наиболее крупные хищники. Высота пирамиды обычно соответствует длине пищевой цепи. И поскольку на верхние этажи пирамиды энергия доходит в очень малых количествах, цепь редко состоит более чем из пяти звеньев.
Ю. Одум сделал расчеты потока энергии от звена к звену в упрощенной теоретической экосистеме, сведя ее к одной примитивной цепи, функционирующей в течение года. Он рассуждал следующим образам. Допустим, имеется посев люцерны на площади в 4 га. На этом поле кормятся телята (предполагается, что они едят только люцерну), а телятиной питается 12-летний мальчик. Результаты расчетов, представленные в виде трех пирамид– численности, биомассы и энергии, свидетельствуют, что люцерна использует всего 0,24 % всей падающей на поле солнечной энергии, из которой 8 % приходится на телят; 0,7 % энергии, накопленной телятами, расходуется на развитие и рост ребенка с 12 до 13 лет. Несмотря на то, что рассматриваемая схема искусственна, она дает четкое представление о масштабах снижения коэффициента полезного действия по мере перехода от основного звена в пирамиде к ее вершине: из всей солнечной энергии, падающей на 4 га люцернового поля, лишь немногим больше миллионной части ее хватает на пропитание мальчика в течение года (рис. 39).
|
|
Дата добавления: 2014-11-06; Просмотров: 3352; Нарушение авторских прав?; Мы поможем в написании вашей работы!