КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Механизм токсического действия
|
|
|
|
Как указывалось выше, действуя в высоких дозах, иприты (сернистый и азотистый) при резорбции нарушают механизмы проведения нервных импульсов в синапсах (главным образом холинергических) центральной нервной системы и на периферии. Этим отчасти объясняются эффекты со стороны сердечно-сосудистой системы (коллапс, брадикардия) или мозга (угнетение высшей нервной деятельности, судороги и т. д.). В последнее время появилась информация о способности ипритов вызывать индукцию и повышать активность NO-синтетазы. Поскольку установлено, что оксид азота является активным регулятором тонуса стенки сосудов и функционального состояния нервных клеток, влиянием на обмен N0 также можно отчасти объяснить развивающиеся сосудистые реакции и нарушения со стороны нервной системы.
Тем не менее основным является цитотоксическое действие ипритов, лежащее в основе большинства патологических процессов, развиваю щихся как на месте аппликации ядов, так и после их поступления во внутренние среды организма. Механизмы цитотоксичности ОВ сложны, многообразны и до конца не выяснены.
Установлено, что на клеточном уровне иприты и активные промежуточные продукты их метаболизма взаимодействуют с нуклеофильными группами молекул клеточных мембран и внутриклеточных структур, вызывая их алкилирование. Основными функционально значимыми мишенями для действия токсикантов являются белки и нуклеиновые кислоты. Взаимодействием с белками можно объяснить ингибиторную активность ипритов в отношении ряда ферментов: гексокиназы, холинацетилазы, ацетилхолинэстеразы, супероксиддисмутазы и т. д. Однако особое значение придают их повреждающему действию на дезоксирибонуклеиновые кислоты (ДНК), формирующие генетический код клетки. В этой связи иприты относят к группе генотоксикантов (веществ, повреждающих генетический код).
В основе повреждающего действия ипритов на ДНК лежит образование ковалентных связей с пуриновыми основаниями нуклеотидов (аде-нином, гуанином).
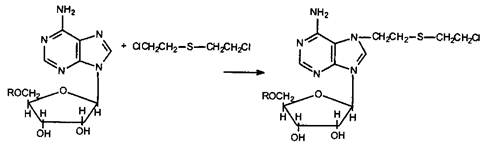
Поскольку иприт обладает двумя функциональными группами, за счет которых осуществляется атака на нуклеофильные группы оснований (рис. 36), возможно «сшивание» комплементарных нитей двойной спирали ДНК (Папирмейстер и соавт. 1993). Уже эта реакция повреждает генетический код клеток, нарушает процессы редупликации и транскрипции, лежащие в основе синтеза белка и клеточного деления. Показано, что иприт блокирует клеточный цикл митоза обратимо в фазе G2M (синтез компонентов клеточных структур, участвующих в процессе деления клеток, например тубулина) и необратимо в фазе GjS (этап утилизации пуриновых и пиримидиновых оснований и синтеза ДНК). Тем не менее алкилирование ДНК является лишь пусковым механизмом процессов, приводящих к еще более глубокому повреждению клеток и их гибели. Как установлено, поврежденные участки ДНК подвергаются депуринизации (отщеплению алкилированных пуриновых оснований от молекулы), а затем депуринизированные участки под влиянием эндонуклеаз «вырезаются» из структуры нитей нуклеиновых кислот. Появление в ядре фрагментов ДНК активирует ферменты репарации этих макромолекул и, в частности, поли (аденозиндифосфорибозо)полимеразу (ПАФРП). Этот энзим участвует в синтезе новых фрагментов ДНК и встраивании их на место поврежденных участков. Поскольку при действии ипритов на клетки повреждаются смежные участки комплементарных нитей ДНК, в процессе репарации возможны грубые ошибки. Иными словами, генетический код клетки полностью не восстанавливается. Как известно, субстратом ПАФРП является никотинамидадениндинуклеотид (НАД), активно потребляемый в ходе репаративных процессов. Истощение этого субстрата (in vitro наблюдается уже через 2 ч после воздействия иприта на культуру клеток) сопровождается нарушением энергообеспечения клетки, снижается уровень АТФ. Это в свою очередь приводит к нарушению внутриклеточного обмена кальция. По данным Гросса и Смитта (1993), концентрация Са2+ в клетках, обработанных ипритом, резко увеличивается, что является пусковым механизмом каскада патологических реакций, приводящих поврежденную клетку к гибели.
В эксперименте показано, что добавление к культуре лимфоцитов ингибиторов поли (аденозиндифосфорибозо)полимеразы (никотинамида, 3-аминобензамида) повышает резистентность клеток к иприту.
Представленные сведения объясняют, почему наибольшей чувствительностью к ипритам обладают органы и ткани, клетки которых активно размножаются (клетки эпидермиса, эпителия желудочно-кишечного тракта, костного мозга и т. д.). Именно здесь нуклеиновый обмен идет с наивысшей интенсивностью, а повреждение генетического аппарата быстро приводит к пагубным последствиям: приостанавливается процесс пополнения пула зрелых, функционально полноценных клеток, выполняющих барьерные, трофические, транспортные и иные функции.
Механизм цитотоксического действия ипритов тесно связан с метаболизмом ксенобиотика в клетках. Полагают, что в реакцию алкилирования биологических субстратов (в том числе и ДНК) вступает не сам иприт, а активные промежуточные продукты его метаболизма. Образование активных метаболитов, как указывалось, проходит при участии микросомальных монооксигеназ. Во второй фазе биопревращения иприта реактивные метаболиты вступают в реакцию конъюгации с глутатионом и деток-сицируются. Такой характер превращения токсиканта создает условия для инициации свободнорадикальных процессов в клетке, во-первых, за счет активации перекисных процессов и, во-вторых, за счет подавления механизмов антирадикальной защиты. Значительное снижение уровня глутатиона в клетках после воздействия иприта и активация в них пере-кисного окисления липидов показаны в эксперименте (Уитфилд, 1987). Активация свободнорадикальных процессов — важный механизм повреждения клеток ксенобиотиками (см. 5.2. «Общие механизмы цитоток-сичности»).
Результатом цитотоксического действия ипритов является инициация ряда патохимических процессов, играющих существенную роль в патогенезе интоксикации. Так, установлено, что под влиянием этих ядов нарушается обмен «медиаторов» воспалительной реакции — цитокинов (эндогенных регуляторов клеточного роста и активности), о чем свидетель ствует изменение их уровня в крови и пораженных тканях. Имеются данные о снижении под влиянием иприта продукции интерлейкина-1о.(IL-la) и увеличении продукции IL-6, IL-8. Продукция интерлейкина-1 f и фактора некроза опухоли (TNF-a) не изменяется. Дисбаланс в продукции цитокинов может существенно влиять на процесс развития воспалительной реакции, вызванной ипритами. Этим, вероятно, можно объяснить вялость течения патологических изменений, скудость клеточных реакций, слабость репаративных механизмов.
Таким образом, имеющиеся в настоящее время данные свидетельствуют о чрезвычайно сложном механизме действия ипритов на организм, выяснение которого очень важно для понимания явления цитотоксично-сти в целом.
|
|
|
|
|
Дата добавления: 2014-01-15; Просмотров: 561; Нарушение авторских прав?; Мы поможем в написании вашей работы!