КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть 8. Физиология редких видов чувствительности
|
|
|
|
Часть 7. Физиология боли.
Часть 6. Зрительный анализатор.
Часть 5. Обоняние у позвоночных.
Часть 4. Вкус у млекопитающих.
Часть 3. Осязание.
Часть 2. Анализа вестибулярной и звуковой информации в мозге.
ТЕМА 12. ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
Часть 1. Равновесие и слух: функция волосковых клеток.
- Структура и функции волосковых клеток.
- Каналы боковой линии.
- Эволюция уха.
- Равновесие.
- Фонорецепция.
- Анатомия и физиология улитки млекопитающих.
- Биофизика волосковых клеток улитки.
- Вестибулярные пути и рефлексы.
- Слуховой путь.
- Слуховая кора.
- Слуховая кора человека и язык.
- Латерализация и нейроанатомия речи.
- Тактильные рецепторы кожи млекопитающих.
- Анализ тактильной информации в мозге.
- Пластичность соматосенсорной коры.
- Строение вкусовых почек.
- Клеточная структура вкусовой почки
- Трансдукция сигнала во вкусовых почках.
- Центральные проекции органа вкуса.
- Распознавание мозгом вкусовых стимулов
- Обонятельные эпителий полости носа
- Обонятельные рецепторные клетки
- Особенности биохимические реакций при передаче сигнала по обонятельным рецепторным клеткам
- Центральная обработка обонятельной информации
- Вомероназальный орган позвоночных и феромоны
- Анатомия глаза
- Особенности строения сетчатки глаза
- Зрительные пути к мозгу
- Первичная зрительная кора
- Экстрастриатная кора
- Распознавание лица
- Прозопагнозия
- Зрительные системы некоторых позвоночных
- Биологическое значение боли
- Нейрофизиология боли
- Иррадиирующая боль
- Воротная теория
- Инфракрасное излучение
- Поляризованный свет
- Электрические поля
- Магнитные поля
Часть 1. Равновесие и слух: функция волосковых клеток.
- Структура и функции волосковых клеток.
 Из всех тонких устройств органов чувств, развитых позвоночными, волосковые клетки – возможно, наиболее удивительные, поскольку они мало отличаются у рыб и у людей. Рассчитано, что на пределе чувствительности уха млекопитающих волосковые клетки стимулируются сдвигом кончика реснички всего на десяток нанометров. Действительно, чувствительность волосковых клеток ограничена только случайным шумом, производимым броуновским движением.
Из всех тонких устройств органов чувств, развитых позвоночными, волосковые клетки – возможно, наиболее удивительные, поскольку они мало отличаются у рыб и у людей. Рассчитано, что на пределе чувствительности уха млекопитающих волосковые клетки стимулируются сдвигом кончика реснички всего на десяток нанометров. Действительно, чувствительность волосковых клеток ограничена только случайным шумом, производимым броуновским движением.
Структура волосковых клеток схематично показана на рис. 1. Из кутикулярной пластинки на дистальном конце клетки вырастает группа до 60 «волосков» двух типов:
ü стереоцилии
ü одиночная киноцилия.
50-60 стереоцилий – это окруженные оболочкой пучки микрофиламентов, а единственная длинная киноцилия – это истинная неподвижная ресничка, часто с булавовидным кончиком. Как показано на рисунке, все волоски сужаются к тонкой «шейке», где они входят в кутикулярную пластинку, а их размер увеличивается по направлению к киноцилии.
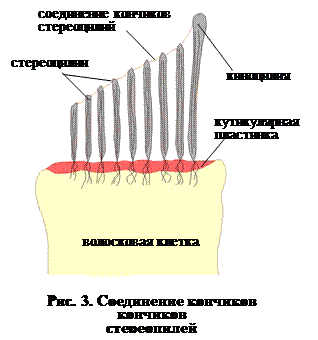 Если рассмотреть стереоцилию при большом увеличении, мы увидим, что она намного длиннее большинства ресничек – 5 мкм, а у ящерицы — даже 30 мкм. У них больше и диаметр – до 900 нм, сужающийся, к основанию до 100 нм. Помимо актиновых микрофиламентов стереоцилии содержат большое количество миозина и кальмодулина. Электронно-микроскопические исследования показывают, что кончики стереоцилий связаны между собой тонкими полипептидными нитями (рис. 2). Они начинаются с клубочка на самой короткой стереоцилий и идут к более длинной соседней. Связывающие стереоцилии волокна находятся под некоторым натяжением, так что, если они повреждаются, мембрана стереоцилий, из которой они исходят, расслабляется.
Если рассмотреть стереоцилию при большом увеличении, мы увидим, что она намного длиннее большинства ресничек – 5 мкм, а у ящерицы — даже 30 мкм. У них больше и диаметр – до 900 нм, сужающийся, к основанию до 100 нм. Помимо актиновых микрофиламентов стереоцилии содержат большое количество миозина и кальмодулина. Электронно-микроскопические исследования показывают, что кончики стереоцилий связаны между собой тонкими полипептидными нитями (рис. 2). Они начинаются с клубочка на самой короткой стереоцилий и идут к более длинной соседней. Связывающие стереоцилии волокна находятся под некоторым натяжением, так что, если они повреждаются, мембрана стереоцилий, из которой они исходят, расслабляется.
Волосковые клетки чрезвычайно чувствительны к механическим стимулам, в чем можно удостовериться, двигая стереоцилию тонким микрозондом в ту и другую сторону. Стереоцилия двигается как жесткое тело – вращаясь вокруг своего суженого основания. Пороговые сдвиги вызывают на мембране волосковой клетки рецепторный потенциал амплитудой около 100 мкВ. Знак этого потенциала зависит от направления движения стереоцилий относительно киноцилии (рис. 3). Если пучок стереоцилий наклоняется по направлению к киноцилии, наблюдается деполяризация; если в противоположную сторону – гиперполяризация. Движение под прямым углом к этой оси не влияет на мембранный потенциал. Эти рецепторные потенциалы достигаются при сдвигах, превышающих ок. 100 нм.

Процесс передачи сигнала очень быстр. Многие млекопитающие, например представители китообразных и рукокрылых, реагируют на звуковые частоты до 100 кГц. В этом случае не хватает времени на биохимические процессы рецепторных взаимодействий и формирование вторичных мессенджеров, которые принимают участие в хемо- и фоторецепции. Воротные механизмы каналов мембраны волосковых клеток должны открываться и закрываться очень быстро. Электрический ответ должен наступать в результате почти мгновенного потока ионов по градиенту концентрации.
Воротные механизмы, регулирующие открывание и закрывание мембранных каналов, представлены в нервной системе двумя основными типами:
Ø первый тип канала управляется электрически, т.е. канал открывается и закрывается в ответ на изменения потенциала клеточной мембраны;
Ø второй тип каналов управляется химически, т.е. открываются,когда особая молекула (медиатор) связывается с рецепторной областью на белке канала. Такие каналы способны реагировать на изменение потенциала на мембране, но очень слабо.
Было установлено, что наибольший ионный поток в ответ на сдвиг наблюдается вблизи от кончика стереоцилий. Ответ, развивающийся очень быстро, начинается через несколько микросекунд после включения стимула, а насыщения достигает приблизительно за 100 мкс.
Проводимость достаточно высока, главным образом для ионов К+, которые присутствуют в эндолимфе и в которую погружены стереоцилии. То обстоятельство, что через канал способны проходить и другие катионы, включая Са2+ и даже малые органические ионы, такие как холин, показывает, что канал имеет достаточно широкую пору – вероятно, ок. 0,7 нм в диаметре. Расчеты показали, что на каждой стереоцилий порядка четырех таких каналов.
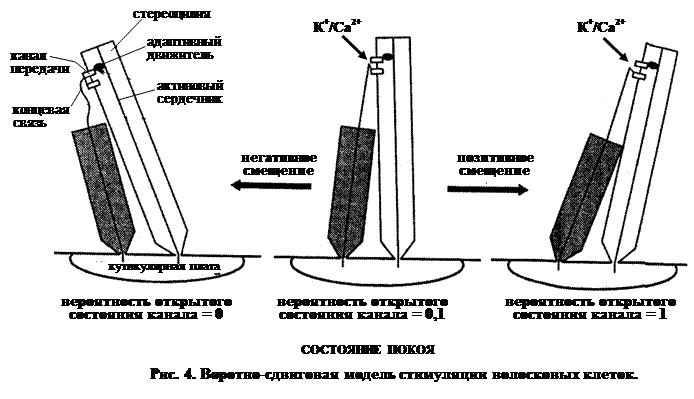
Биофизическая модель стереоцилий была предложена Хадспетом. Как было отмечено выше, каждая стереоцилия связана вблизи от кончика молекулярной нитью (концевой связью) с соседней более длинной (рис. 2). Накопились данные, показывающие, что эти нити присоединены к ионным каналам. Предполагается, что, когда стереоцилия находится в покое, наблюдается небольшая утечка через каналы. Как показано на рис. 4, вероятность открытого состояния канала в таком случае составляет около 0,1.
Когда механический стимул вызывает сдвиг группы стереоцилий в сторону киноцилии, у более длинных стереоцилий вероятность открытия каналов сдвигается к единице. Поскольку реснички погружены в богатую ионами калия среду (эндолимфу), то К+ является основным ионом, входящим по градиенту концентрации в ресничку, что вместе с потоком Са2+ в клетку вызывает деполяризацию мембраны.
Если стереоцилий сдвигаются противоположном направлении, вероятность открывания каналов смещается к нулю. Поток К+/Са2+ прекращается и мембрана гиперполяризуется.
 Описанный выше механизм делает возможным почти мгновенный ответ на механическую стимуляцию (что и наблюдается). В то же время известно, что большинство волосковых клеток очень быстро адаптируется (за несколько десятков миллисекунд).
Описанный выше механизм делает возможным почти мгновенный ответ на механическую стимуляцию (что и наблюдается). В то же время известно, что большинство волосковых клеток очень быстро адаптируется (за несколько десятков миллисекунд).
Иными словами, когда отклонение стереоцилий поддерживается более миллисекунды, поток К+/Са2+ внутрь клетки и последующая деполяризация прекращаются.
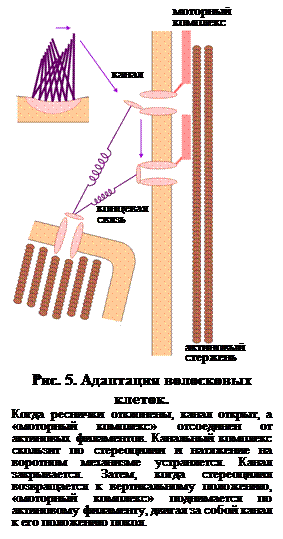
Это становится возможным благодаря наличию следующего механизма в стереоцилиях. Схема его ультраструктуры показана на рис. 5. Из рисунка видно, что канальный белок, помимо концевой связи, соединен через миозиновый «движитель» с внутренними актиновыми микрофиламентами. Это соединение обеспечивает основу для стабилизации положения канала в мембране, так что усиление натяжения на концевой связи не сдвигает весь канальный белок по стереоцилии в плоскости мембраны.
Механизм, ответственный за адаптацию, зависит от потока Са2+ внутрь клетки при открытом канале. Одна из теорий предполагает, что это приводит к разрыву связи между миозиновым «движителем» и актиновыми микрофиламентами в стереоцилии. Это в свою очередь позволяет ионному каналу скользить по мембране стереоцилий, устраняя тем самым натяжение концевой связи. Вероятность открытого состояния канала снижается и поляризация мембраны возвращается к уровню покоя. Когда стереоцилия возвращается к нормальному неизогнутому состоянию, вероятность открытого состояния канального белка, который теперь не испытывает натяжения, приближается к нулю. В отсутствии потока Са2+ миозиновый движитель тянет канал по актиновому филаменту, возвращая натяжение концевой связи к исходному значению и вероятности открытого состояния канала к 0,1.
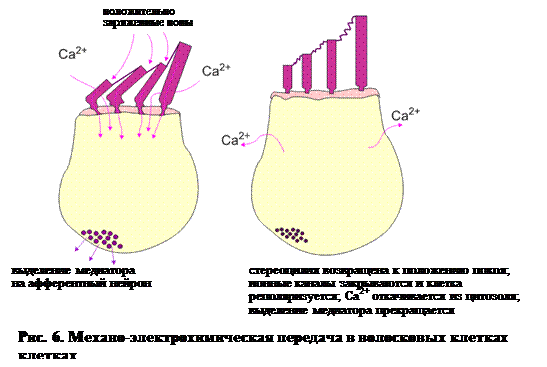 Помимо потока Са2+ через трансдукционные каналы ресничек, ионы Са2+ входят также в клетку через потенциалзависимые каналы в кутикулярной пластинке. Эти каналы открываются, когда мембрана деполяризуется вследствие отклонения стереоцилии. Поток Са2+ приводит в свою очередь к выделению медиатора из основания волосковой клетки (рис. 6). В зависимости от природы медиатора и природы подлежащего дендрита это инициирует или ингибирует синаптический потенциал. В первом случае запускается потенциал действия, который распространяется по сенсорному нерву в центральную нервную систему, во втором – гиперполяризация.
Помимо потока Са2+ через трансдукционные каналы ресничек, ионы Са2+ входят также в клетку через потенциалзависимые каналы в кутикулярной пластинке. Эти каналы открываются, когда мембрана деполяризуется вследствие отклонения стереоцилии. Поток Са2+ приводит в свою очередь к выделению медиатора из основания волосковой клетки (рис. 6). В зависимости от природы медиатора и природы подлежащего дендрита это инициирует или ингибирует синаптический потенциал. В первом случае запускается потенциал действия, который распространяется по сенсорному нерву в центральную нервную систему, во втором – гиперполяризация.
Волосковые клетки эволюционно очень древние и развились во всех классах позвоночных – от бесчелюстных до млекопитающих. У круглоротых с их двумя современными представителями, миксиной и миногой, волосковые клетки развились и в каналах боковой линии, и в ухе.
|
|
|
|
Дата добавления: 2014-01-15; Просмотров: 374; Нарушение авторских прав?; Мы поможем в написании вашей работы!