КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Основы эмбриогенеза человека. Не прибегая к разложению в ряд Фурье, определить коэффициенты амплитуды и формы кривой на рис
|
|
|
|
Не прибегая к разложению в ряд Фурье, определить коэффициенты амплитуды и формы кривой на рис. 4.
Ответ: 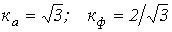 .
.
7. Определить действующее значение напряжения на зажимах ветви с последовательным соединением резистора с 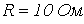 и катушки индуктивности с
и катушки индуктивности с 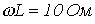 , если ток в ней
, если ток в ней 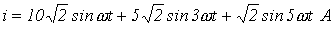 . Рассчитать активную мощность в ветви.
. Рассчитать активную мощность в ветви.
Ответ: U=218 В; Р=1260 Вт.
8. Определить действующее значение тока в ветви с источником ЭДС в схеме на рис. 5, если 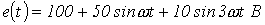 ;
; 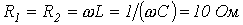 .
.
Ответ: I=5,5 A.
Эмбриогенезу человека предшествует прогенез – процессы развития половых клеток, т.е., овогенез и сперматогенез. Первые 3 стадии развития половых клеток схожи и в тоже время имеют различия:
I – стадия размножения: происходит размножение стволовых половых клеток путем деления митозом малодифференцированных половых клеток (светлые сперматогонии типа А и овогонии). Стадия размножения у лиц женского пола протекает еще в эмбриональном периоде, у лиц мужского пола – после полового созревания.
II – стадия роста: происходит увеличение размеров половых клеток и подготовка к мейозу (происходит в стадии созревания) – синтез ДНК и перекомбинация генов в гомологичных хромосомах при кроссенговере. У лиц мужского пола происходит после полового созревания и при этом сперматогонии превращаются в сперматоциты I порядка, у лиц женского пола эта стадия протекает в 2 периода: “период малого роста” – в эмбриональном периоде происходит незначительное увеличение размеров половой клетки, синтез ДНК и перекомбинация генов в гомологичных хромосомах при кроссенговере, “период большого роста” – после полового созревания, при этом овоцит I порядка почти в 2 раза увеличивается в размерах, приобретает вторичную оболочку, накапливает желток (трофические включения).
III – стадия созревания: происходит мейоз (2 быстро следующих друг за другом деления половых клеток без удвоения хромосом), в результате образуются половые клетки с уже с гаплоидным набором хромосом. У мужчин стадия созревания происходит после полового созревания, а у лиц женского пола – начинается еще в эмбриональном периоде и завершается после полового созревания. У мужчин в стадии созревания из одного сперматоцита I порядка образуется 4 сперматида; у женщин первое деление начинается еще в эмбриональном периоде, но остается незавершенным, продолжается и заканчивается после полового созревания и из одного овоцита I порядка образуется один овоцит II порядка и первое редукционное (направительное) тельце, а после второго деления – одна яйцеклетка и второе редукционное тельце. Первое редукционное тельце также может поделиться, поэтому в результате стадии созревания у женщин из одного овоцита I порядка образуется 1 яйцеклетка и 3 (или 2) редукционные тельца.
IV – стадия формирования имеет место только при сперматогенезе, при этом сперматиды избавляются от излишнего количества цитоплазмы, покрываются гликокаликсом, приобретают характерный для них структуру (морфологию), т.е. превращаются в сперматозоиды.
В результате гаметогенеза образуются половые мужские женские клетки, которые отличаются от соматических клеток по следующим признакам:
1. Набор хромосом: соматические клетки имеют диплоидный, половые клетки – гаплоидный набор хромосом.
2. Половые клетки имеют специальные приспособления для выполнения своих специфических функций:
а) сперматозоиды – акрасому (видоизмененный пластинчатый комплекс) для проникновения через оболочки яйцеклетки и двигательный аппарат (хвостик, центриоли и митохондрии);
б) яйцеклетки – I и II оболочки, желток (трофические включения).
3. Ядерно-цитоплазматическое отношение половых клеток резко отличается от соматических: в яйцеклетке – очень низкое (резко преобладает масса цитоплазмы), в сперматазоидах – очень высокое (преобладает масса ядра).
4. Жизненный цикл у половых сложный и многостадийный.
5. Биологическое назначение: из соматических клеток могут образоваться только подобные клетки, а из половых клеток – целый новый организм.
Яйцеклетка человека имеет диаметр около 130 мкм, окружена первичной (оолеммa или собственная оболочка) и вторичной (блестящая или прозрачная зона — Zоnа ро11ucida=Zр) оболочками и слоем фолликулярных клеток. Блестящая зона состоит из гликопротеинов Zр1, Zр2 и ZрЗ (является специфическим рецептором для сперматозоидов) и гликозаминогликанов. Ядро содержит гаплоидный набор хромосом и хорошо выраженное ядрышко. В цитоплазме имеются гранулярный ЭПС, комплекс Гольджи, умеренное количество митохондрий, гранулы желтка (трофические включения), сразу под цитолеммой — кортикальные гранулы (гранулы диаметром около 1мкм. окружены элементарной.мембраной, содержат ферменты для кортикальной реакции). В яйцеклетки человека же’лтка мало, желток распределяется по цитоплазме равномерно, т.е. яйцеклетка олиголецнтальная и изолецитальная.
Процесс внутриутробного развития у человека длится.в среднем 280 суток (10 лунных месяцев) и делится на 3 периода:
1. Начальный период (1-ая неделя).
2. Зародышевый период (2-8 недели).
3. Плодный период (с 9-й недели до рождения).
Эмбриогенез
включает в себя процессы с момента оплодотворения до рождения и включает следующие его дни.
1. Оплодотворение, в результате которого образуется зигота (одноклеточный зародыш),
2. Дробление зародыша с образованием бластулы.
3. Гаструляция — образование 3-х листкового зародыша.
4. Гистогенез, органогенез и ситемагенез — дифференцировка зародышевых листков в ткани органов, образование из органов систем органов.
Оплодотворение — это сближение и слияние половых клеток с образованием одноклеточного зародыша — зиготы. У человека оплодотворение внутреннее, т.е. происходит и женских половых путях. В процессе оплодотворения выделяют:
1. Дистантное взаимодействие и сближение половых клеток.
2. Контактное взаимодействие половых клеток и активизация яйцеклетки.
3. Вхождение сперматазоида в в яйцеклетку и последующей синкарион (сингамия)- слияние женского и мужского пронуклеусов.
Дистантное взаимодействие (взаимодействие на расстоянии) половых клеток начинается с момента попадания сперматозоидов в женские половые пути, т.е. когда сперматозоиды находятся еще во влагалище: женские половые клетки выделяют специфические вещества — гемогомоны, которые вызывают хемотаксис сперматозоидов — свойство сперматозоидов двигаться против градиента концентрации (туда, где выше концентрация) гемогомонов Хемотаксис сперматозоидов обуславливает их направленное движение к яйцеклетке. Продвижению сперматозоидов к яйцеклетке способствует также реотаксис сперматозоидов — свойство сперматозоидов всегда двигаться против тока жидкосж (слизь в женских половых путях течет по направлению: маточные трубы —> матка —> влагалище).
Сперматозоиды двигаются со скоростью 30-50 мкм/сек. Поэгому уже через 30-60 минуг могут попадать в полость матки, а через 1,5-2 часа — ампулярной части маточной трубы. Сразу после эякуляции и в. ближайшие часы сперматозоиды еще не способны оплодотворить яйцеклетку, для этого должна произойти капацнтация сперматозоида -приобретение способности к оплодотворению под воздействием секретов женских половых путей. Для капацитации сперматозоидов в среднем требуется 7 часов.
Контактное взаимодействие половых клеток и проникновение сперматозойда в яйцеклетку Многочисленные сперматозоиды окружают яйцеклетку и выделяют ферменты, под воздействием которых о г яйцеклетки отсоединяются фолликулярные клетки. Сперматозоиды синхронным биением жгутиков вращают яйцеклетку со скоростью 4 оборота в минуту (в течении до 12 часов!). При помощи специфических (строго комплементарных для каждого вида) рецепторов сперматозоид (фермент гликозилтрансфераза па повнерхностп акросомы) и яйцеклетка (сахар N-ацетилглюкозамин в составе ZрЗ фракции гликопротеинов блестящей зоны) «узнают» друг друга — т.е. происходит контакт, в результате у сперматозоида просходит акрасомальная реакция — выделение и воздействие на оболочки яйцеклетки ферментов акросомы (гиалуронидаза и трипсин) и головка сперматозоида проникает через блестящую зону и оолемму и цитоплазму яйцеклетки. У человека и яйцеклетку проникает только один сперматозоид — моноспермия. Проникновению других сперматозоидов (полиспермия) препятствует наступающая через» несколько секунд кортикальная реакция — мембраны кортикальных гранул яйцеклетки сливаются с оолеммой и содержимое гранул воздействует на блестящшую зону (И оболочку яйцеклетки); в результате этого блестящая оболочка утрачивает способность осуществлять специфический контакт со сперматозоидами и запускать у них акрасомальную реакцию, уплотняется и превращается в оболочку оплодотворения, непроницаемую для остальных сперматозоидов.
Синкарион. После проникновения в яйцеклетку ядро сперматозоида увеличивается в размерах, хроматин десперализуется. Далее ядра яйцеклетки и сперматозоида (пронуклеусы) сближаются и сливаются (синкарион), а из клеточного цетра сперматозоида начинает формироваться веретено деления — в результате образуется зигота, т.е. одноклеточный зародыш. В зиготе начинается интенсивный синтез белка. — яйцеклетка актиивизируется, начинает готовится к следующей стадии эмбриогенезе — дроблению. Для активизации яйцеклетки проникновение в нее сперматозоида необязательно, достаточно кратковременного контакта между поверхностью яйцеклетки и сперматозоида. Яйцеклетку можно активизировать с помощью множества неспецифических химических и физических факторов (например укол иглы). Развитие яйцеклетки активацией без сперматозоида называется партеногенезом (пример: у некоторых насекомых, у пчел).
В настоящее время разработаны методы оплодотворения яйцеклеток млекопитающих и человека in vitro — в пробирке, с последующей пересадкой в матку. Благодаря этой методике бесплодные женщины получили возможность рожать нормальных детей.
Дробление — это деление зародыша, но это деление несколько отличается от обычного деления митозом: при дроблении отсутствует пресинтетнческая фаза G1, поэтому дочерние (слетки (бластомеры) после митоза не синтезируют белки и не растут — с каждым делением размеры образовавшихся бластомеров становится все меньше и меньше (отсюда и название «‘дробление»). В результате дробления восстанавливается нормальное ядерно-цитоплазматическое отношение (у оплодотворенной яйцеклетки оно очень низкое) и образуется бластула. Тип дробления зависит от количества и распределения желтка в яйцеклетке. Олиголецитальная изолецитальная яйцеклетка человека дробится по типу: полное неравномерное асинхронное. Полное — в дроблении участвуют все участки оплодотворенной яйцеклетки; неравномерное — образовавшиеся бластомеры не одинаковые, не равные: одни крупные и темные, располагаются в центре зародыша их совокупность называется змбриобластом — будущее тело, другие мелкие и светлые, окружают снаружи бластомеры эмбриобласта — их совокупность называется трофобластом — участвует при формировании плаценты; асинхронное = количество бластомеров увеличивается не по геометрической прогреcии (2-> 4-> 8 и т.д., т.е. кратное увеличение числа блаcтомеров). В то же время следует отметить, что первые бластомеры (по крайней мере до 8 первых бластомеров) потенциально не различаются. Доказательством этого утверждения является образование однояйцевых (идентичных) близнецов. Однояйцевые близнецы образуются из одной яйцеклетки в случае когда по каким-то причинам на стадии 2-х или 3-х бластомерного зародыша происходит их разделение и из каждого бластомера развивается самостоятельный отдельный организм. С другой стороны, если в эксперименте взять двухбластомерного зародыша и разрушить, один бластомер, то из второго бластомера может развиться вполне нормальный организм. Или еще; если взять двух эмбрионов на стадии до 8-бласгомерного этапа и их объединить в одну морулу, то можно получить новый организм, наследующий признаки не 2-х а 4-х родителей — такие экспериментально полученные животные называются химерами. Этими экспериментами было доказано, что бластомеры ранних стадий дробления (вплоть до 8-клеточной стадии) практически идентичны и обладают неограниченными потенциями. С помощью метода химер было доказано, что какие бластомеры дадут трофобласт, а какие — эмбриобласт зависит не от особенностей самих бластомеров, а лишь от места, какое они случайно заняли в моруле. Бластомеры расположенные снаружи морулы подвергаются воздействию одних условий, внутренние бластомеры оказываются в других условиях, что и определяет направление их дальнейшей дифференцировки.
Дробление зиготы человека начинается в конце 1-х суток после оплодотворения в дистальной части маточной трубы и заканчивается в 7-е сутки в полости матки. На 2-3-и сутки зародыш находится в маточных трубах и имеет вид плотного узелка — морулы, в центральной части которого находятся крупные темные бластомеры — эмбриоблзст, а по периферии -мелкие светлые бластомеры = трофобласт. В 4-е сутки бластула находится в проксимальной части маточной трубы, т.е. вплотную подходит к полости матки и имеет вид пузырька. Бластомеры трофобласта всасывают секрет маточной трубы и сами секретируют жидкость, поэтому трофобласт растягивается и превращается в пузырек, заполненный жидкостью, а эмбриооласт прикрепляется на одном полюсе к трофобласту снутри. Такая бластула называется эпибластулой (или синонимы; бластоциста, стерробластула). На 5-е сутки бластоциста попадает в полость матки и остается там до 7-х суток, за это время несколько увеличивается в размерах (100 и более бластомеров).
Гаструляция — это сложный процесс, где в результате размножения, роста, днфференцировкн н направленного перемещения клеток образуется 3-х листковый зародыш. Гаструляция происходит 7-17 сутки и осуществляется путем деламииацин или расщипления (7-14 сутки) и иммиграции или выселения (14-17 сутки).
В 7-е сутки эмбриобласт расщепляется на 2 слоя: верхний слой — эпибласт или первичная эктодерма (содержит материал будущей эктодермы, мезодермы, хорды и части энтодермы) и нижний слой — гипобласт (будущая энтодерма после присоединения клеточного материала прехордальной пластинки из эпибласта). Почти одновременно с этим происходит выселение клеток из эпи- и гипооласта — внезародышевая мезенхима, которая выстилает внутреннюю поверхность трофобласта. Далее в течение 2-й недели эпибласт и гипобласт начинают прогибаться в противоположных направлениях и превращаются в пузырьки: из эпибласта образуется амниотнческий пузырек, нз гипобласта — желточный пузырек. Эти 2 пузырька окружаются внезародышевой мезенхимой. Соприкасающиеся поверхности амниотнческого и желточного пузырька имеют вид диска (или щитка) и соответственно называются зародышевым эпибластом и зародышевым гипобластом, а вместе — зародышевым щитком. Остальные участки амниогического и желточного пузырька называются внезародышевым эпи- и гипобластом.
В начале 3-й недели (14-17 сутки) происходит иммиграция (выселение) клеток из эпибласта, причем это происходит в 2 фазы: в I фазе идет подготовка к выселению — клеточный материал подлежащий иммиграции перемещается (медленно двигающиеся клетки: с будущего краниального конца к каудальному концу по центру эпибласта, а быстродвигаюшиеся клетки: вначале тоже с краниального полюса к каудальному полюсу, но по краю э:.’нблпста, а затем поворачиваются и идут но центру эпибласта к краниальному концу) и собирается вместе и образует на поверхности эпибласта 3 структуры: прехордальную пластинку, I узелок и I полоску; II фаза — собственно выселение материала этих 3- структур. I узелок выселяется и между имеющимися двумя листками образует первый осевой орган — хорду. Материал прехордальной пластинки выселяется и присоединяется к гипобласту, с этого момента нижним листок будет называться энтодермой.
Клетки 1 полоски выселяется, занимает положение между двумя имеющимися листками и образует средний листок — мезодерму. Оставшаяся часть эпибласта после выселения клеток 3-х структур будет называться эктодермой.
В следующей стадии начинается дифференцировка зародышевых листков в ткани (гистогенез) органов (органогенез) и формирование из органов систем органов (системогенез). При этом следует выделить 20-21 сутки эмбриогенеза — в эти сроки происходят следующие важные процессы:
1 Из всех трех зародышевых листков, но преимущественно нз мезодермы, выселяются клетки, заполняют пространства между тремя зародышевыми листками, т.е. формируется зародышевая мезенхима.
2. Мезодерма дифференцируется на составные части (томы) сомиты, сегментные ножки и спланхнотомы.
3 Трехлистковый плоским зародыш сворачивается «в трубку» — формируется туловище (энтодерма сворачиваясь в трубку превращается к I кишку, окруженную слоем мезенхимы и сплапхнатомами; эктодерма покрывает туловище снаружи).
4. При сворачивании зародыша» в трубку» внезародышевые части организма обособляются от тела зародыша и формируются провизорные органы желточный мешок, гипнотическая оболочка.
Материал зародышевых листков дифференнцируется в ткани, органы и системы органов (более подробно о развитии конкретных органов и систем смотри в лекциях по частной гистологии):
I Эктодерма:
- нервная ткань органов нервной системы;
- эпидермис кожи и его производные (сальные, потовые, молочные железы, ногти, волосы);
- эпителии роговицы и хрусталик глаза, эпителий преддверья ротовой полости и анального отдела прямой кишки;
II Мезодерма: -дерматомы -> дерма кожи;
- миотомы —> скелетная мускулатура;
- склеротомы —> осевой скелет (кости и хрящи позвоночного столба;
- сегментные ножки (нефрототомы) —► эпителий мочеполовой системы;
- спланхиотомы -> мезотелий серозных покровов (брюшины, плевры и околосердечной сумки), эпителий гонад (клетки Сертоли яичка и фолликулярные клетки яичников), корковая часть надпочечников, миокард и эпикард;
III Энтодерма:
- эпителий и железы (включая печень и поджелудочную железу) пищеварительном и дыхательной системы;
IV Мезенхима:
ткани внутренней среды (кровь и лимфа, все виды волокнистых соединительных тканей — рыхлая волокнистая соединительная ткань, плотная волокнистая оформленная и неоформленная волокнистая соединительные ткани, соединительные ткани со специальными свойствами, костные и хрящевые ткани) и гладкая мышечная ткань.
Начало формы
Основы сравнительной эмбриологии
Эмбриология — это наука о закономерностях эмбрионального развития организма от момента оплодотворения до рождения. Нас в большей степени интересует медицинская эмбриология, которая изучает закономерности развития зародыша человека, причины уродств и пути влияния на эмбриональное развитие. Но сегодня мы рассмотрим эмбриональное развитие в сравнительном аспекте у ланцетника, лягушки, птиц и млекопитающих, т.к. у человека наблюдается явление рекапитуляции — т.е. повторение многих этапов эмбрионального развития этих видов.
Методы эмбриологических исследований:
1. Визуальное наблюдение развития зародышей, настоящее время дополнительно фиксируемое микрокино- или видеосьемками.
2. Метод изучения фиксированных зародышей на разных этапах с последующей микроскопией.
3. Метод маркировки клеток с последующим прослеживанием перемещений маркированных клеток в тканях и органах зародыша. В качестве маркера раньше использовали угольную пыль, позже — нейтральные красители, в настоящее время используют антитела к определенным белкам развивающегося зародыша, причем эти антитела обычно мечены флуоресцином.
4. Метод микрохирургии — удаление отдельных частей зародыша.
5. Метод трансплантации части от одного зародыша к другому.
Началу индивидуального развития предшествует возникновение половых клеток, т.е. гаметогенез, который можно рассматривать прогенезом при индивидуальном развитии. Более подробно гаметогенез мы будем изучать в теме «Половая система». Поэтому ограничимся перечислением отличий половых клеток от соматических клеток:
Набор хромосом у половых клеток гаплоидный, у соматических — диплоидный.
Для половых клеток характерно сложное, стадийное развитие; при этом имеет место особый способ деления — мейоз.
Половые клетки имеют специальные приспособления:
- сперматозоид имеет акросому (для проникновения через оболочки я/к) и мощный двигательный аппарат — хвостик;
- яйцеклетка имеет желток (запас питательных веществ и строительных материалов) и оболочки (I, II, а у некоторых видов и III).
4. У половых клеток особое ядерно-цитоплазматическое отношение: у мужских пол. клеток очень высокое (преобладает ядро над цитоплазмой), в женских половых клетках очень низкое (преобладает цитоплазма над ядром).
5. Обмен веществ в зрелых половых клетках до оплодотворения находится на очень низком уровне (почти до анабиоза).
6. Биологическое назначение: если из соматической клетки может образоваться лишь такая же дочерняя клетка, то из половых клеток формируется целый новый организм.
Для объяснения течения начальных этапов эмбриогенеза большое значение имеет знание особенностей строения яйцеклетки (я/к), поэтому остановимся на классификации я/к. В основу классификации положены 2 признака: количество и распределение желтка в я/к.
По количеству желтка различают следующие виды я/к:
1. Алецитальные (безжелтковые) — у видов, развитие которых протекает с метаморфозами и эмбриональный период очень короткий или у некоторых паразитарных червей.
2. Олиголецитальный (маложелтковый) — у видов развивающихся вне организма матери в относительно благоприятной водной среде, эмбриональный период относительно короткий (пример: ланцетник). А также у видов развивающихся внутриутробно и питающихся за счет матери (пр.: млекопитающие).
3. Мезолецитальные (среднее количество желтка) — развитие вне организма матери в водной среде (пр.: лягушка).
4. Полилецитальные (многожелтковые) — развитие идет вне организма матери, причем на суше (пр.: птицы, пресмыкающиеся).
ВЫВОД: количество желтка в я/к зависит от условий где развивается зародыш, а также в какой то степени от длительности эмбрионального развития.
Вышеприведенная классификация дополняется классификацией по распределению желтка по цитоплазме:
1. Изолецитальная (равномерное распределение) — характерно для олиголецитальных я/к. Различают I изолецитальную (ланцетник) и II изолецитальную я/к (млекопитающие).
2. Телолецитальные я/к — желток распределяется по цитоплазме неравномерно, полярно — на одном полюсе (вегетативный) желток, а на другом полюсе (анимальный) ядро и органоиды. Характерно для мезо- и полилецитальных я/к; Среди телолецитальных различают 2 подгруппы:
а) умеренно телолецитальные — полярность выражено умеренно, нерезко (пр.: мезолецитальная я/к лягушки);
б) резко телолецитальные — полярность ярко выражена (пр.: птицы).
3. Центролецитальные — желток в виде узкого пояска сосредоточен вокруг ядра.
Итак у ланцетника я/к олиголецитальная I изолецитальная, у лягушки — мезолецитальная умеренно телолецитальная, у птиц — полилецитальная резко телолецитальная, у млекопитающих — олиголецитальная II изолецитальная.
Оболочки я/к: I оболочка — собственная оболочка (оолемма), II оболочка — продукт деятельности самой я/к и соседних вспомогательных клеток (например фолликулярных клеток); III оболочка имеется у видов развивающихся вне организма матери на суше, и является продуктом деятельности слизистой яйцевыводящих путей.
В эмбриогенезе различают следующие этапы:
1. Оплодотворение.
2. Дробление.
3. Гаструляция.
4. Гистогенез, органогенез, системогенез (дальнейшая дифференцировка зародышевых листков).
Оплодотворение бывает наружным (у видов развивающихся в водной среде) и внутренним. При оплодотворении различают:
1) дистантное взаимодействие половых клеток;
2) сближение половых клеток;
3) проникновение мужской половой клетки в женскую.
При дистантном взаимодействии большое значение имеют хемотаксис и реотаксис. Хемотаксис - способность мужских половых клеток двигаться только против градиента концентрации гемогомонов (специфические вещества, выделяемые женской половой клеткой), т.е. мужская половая клетка двигается туда, где выше концентрация гемогомонов. Концентрация гемогомонов выше всего вокруг женской половой клетки, и уменьшается по мере удаления от я/к. Реотаксис — способность спематозоидов двигаться только против тока жидкости. А жидкость в женских половых путях течет: в маточных трубах по направлению к матке, а в матке — по направлению к влагалищу.
Кроме таксисов сближению половых клеток способствуют:
- перистальтика маточных труб;
- мерцательное движение ресничек эпителия маточных труб.
На близком расстоянии встрече половых клеток способствует противоположная заряженность половых клеток. Распознавание половых клеток после контакта осуществляется при помощи специфических рецепторов. После контакта только одна мужская половая клетка при помощи ферментов акросомы проникает в я/к; оболочка я/к изменяет свои свойства, становится непроницаемой для других сперматозоидов, т.е. образуется оболочка оплодотворения.
Дробление - это деление оплодотворенной я/к (уже зародыша) митозом. Дочерние клетки называются бластомерами, они не расходятся. При дроблении очень короткие интерфазы, поэтому бластомеры не успевают расти, а наоборот с каждым делением становятся размерами все меньше и меньше, т.е. количество бластомеров увеличивается, а обьем каждого отдельного бластомера уменьшается. Тип дробления зависит от типа я/к, т.е. от количества и распределения желтка. Характеризуя тип дробления у разных видов нужно ответить на 3 вопроса:
1. Полное (голобластическое) или неполное (меробластическое) дробление.
2. Равномерное или неравномерное дробление.
3. Синхронное или асинхронное дробление.
Полное дробление — когда в дроблении участвуют все участки зародыша; характерно для олиго-изолецитальных(ланцетник, млекопитающие), а также мезо-умеренно телолецитальных я/к (лягушка).
Неполное дробление — когда дробление идет только на анимальном полюсе, вегетативный полюс перегружен желтком и в дроблении не участвует. Характерно для поли- и резко телолецитальных я/к (птицы).
Равномерное дробление — образовавшиеся бластомеры равные, одинаковые; хар-но для олиго- и I изолецитальных я/к (ланцетник).
Неравномерное дробление — образовавшиеся бластомеры неравные, разные: одни крупные, другие мелкие; одни дифференцируются в тело зародыша, другие — для питания; хар-но для мезо- и полилецитальных (лягушка, птица), а также для олигоIIизолецитальных я/к (млекопитающие).
Синхронное дробление — когда все бластомеры дробятся одинаковой скоростью и поэтому количество их увеличивается по правильной прогрессии, т.е. кратное увеличение; как-то: 1 ® 2 ® 4 ® 8 и т.д.
Асинхронное дробление — кол-во бластомеров увеличивается по неправильной прогрессии; как-то: 1 ® 2 ® 3 ® 5 — и т.д.
У ланцетника дробление полное, равномерное, синхронное. В результате такого др-ия у ланцетника образуется целобластула — полый пузырек, заполненный жидкостью. Стенка целобластулы (бластодерма) образована одним слоем бластомеров и в ней различают крышу, дно и краевую зону.
У лягушки дробление полное, неравномерное, асинхронное; в результате образуется амфибластула, состоящая из анимального и вегетативного полюса и бластоцели с жидкостью. Бластомеры анимального полюса мелкие, дифференцируются в последующем в тело зародыша, а бл-меры вегетативного полюса крупные, перегружены желтком и обеспечивают питание зародыша.
У птиц дробление неполное (дискоидальное), неравномерное и асинхронное; в рез-те обра- зуется дискобластула. Желток в др-ии не участвует, остается как одно целое; дроб-ие идет только на анимальном полюсе. т.е. где ядро и органоиды я/к. Образовавшиеся бл-меры распластываются на желтке и наз-ся зародышевым щитком; между зародыш. щитком и желт- ком имеется узкая щель — бластоцель.
У млекопитающих дробление полное, неравномерное, асинхронное; в рез-те образуются бл-меры 2-х типов: в центре крупные темные бл-меры — это эмбриобласт, дифф-ся в тело; по периферии мелкие светлые бл-меры — это трофобласт, участвующий при формировании хориона и плаценты. Вначале образуется морула (полости еще нет), впоследствии трофобласт всасывает жидкость слизистой яйцевыводящих путей, поэтому морула превращается в полый пузырек — эпибластула (синоним — стерробластула): стенка пузырька из одного слоя бластомеров трофобласта; полость (бластоцель) пузырька заполнена жидкостью; на одном полюсе к трофобласту изнутри прикреплен эмбриобласт.
После дробления начинается следующий этап — гаструляция. Гаструляция — это сложный процесс, где в результате размножения, роста, дифференцировки и направленного перемещения бластомеров образуется трехлистковый зародыш, т.е. образуются зародышевые листки: эктодерма, энтодерма и мезодерма.
У ланцетника гаструляция происходит способом инвагинации (впячивание): днобластулы постепенно впячивается под крышу и формируется эктодерма и энтодерма; при этом образуется гастроцель и гастропора. Мезодерма образуется путем выпячивания энтодермы.
У лягушки гаструляция происходит способом эпиболии (обрастание): бластомеры анимального полюса делятся быстрее и начинают обрастать вегетативный полюс.
У птиц гас-ия очень похожа с гас-ией у млекопитающих, поэтому нужно хорошо разобраться. Гаст-ия идет в 2 этапа:
I этап деламинация (расщепление), II этап — иммиграция (выселение). На I этапе зародышевый щиток расщепляется на 2 листка: верхний — эпибласт, нижний — гипобласт.
II этап -иммиграция, состоит из 2-х фаз: I фаза — подготовка к иммиграции, в рез-те образуются на поверхности эпибласта прехордальная пластинка, I узелок и I полоска:
Оставшаяся часть эпибласта после выселения клеток прехордальной пластинки, I узелка и I полоски называется эктодермой. Гипобласт после присоединения к нему клеток прехордальной пластинки называется энтодермой. Клетки I узелка выселяясь образуют первый осевой орган — хорду, а I полоска выселяясь образует мезодерму.
После гаструляции начинается следующий этап эмбрионального развития — дальнейшая дифференцировка зародышевых листков с образованием из них тканей, органов и систем органов (гистогенез, органогенез, системогенез).
Мезодерма подразделяется на 3 части: дорсальная часть — сомиты, которые в свою очередь состоят из дерматомов, миотомов и склеротомов; вентральная часть мезодермы — спланхнотомы, состоящие из париетальных и висцеральных листков; часть мезодермы соединяющая сомиты со спланхнотомами в передней части туловища сегментируется и назвается нефрогонотомами (синоним: сегментные ножки), а в задней части туловища не сегментируется и называется нефрогенной тканью.
Пространство между 3-мя зародышевыми листками заполняется мезенхимой (образуется путем выселения из всех 3-х листков, но преимущественно из мезодермы).
Из эктодермы в дорсальной части путем впячивания образуется еще один осевой орган — нервная трубка, из которой потом образуется вся нервная система.
Гаструляция у млекопитающих протекает в принципе аналогично у птиц, хотя имеются некоторые особенности. На I стадии путем деляминации из эмбриобласта образуются также эпибласт и гипобласт. Дальше эпибласт и гипобласт начинают прогибаться в противоположных направлениях и образуют соответственно 2 пузырька: из эпибласта — амниотический, из гипобласта — желточный. Лишь только после этого начинается II этап гаструляции — иммиграция, протекающая практически также как у птиц.
II этап гаструляции — иммиграция начинается на части эпибласта, являющейся дном амниотического пузырька: I фаза — подготовка к выселению с образованием на поверхности дна амниотического пузырька прехордальной пластинки, I узелка, I полоски. А дальше идет II фаза иммиграции — собственно выселение клеток этих 3-х структур: клетки прехордальной пластинки включаются в состав гипобласта и образуется энтодерма; из I узелка образуется хорда, а из клеток I полоски после выселения образуется средний зародышевый листок — мезодерма.
После гаструляции начинается дальнейшяя дифференцировка зародышевых листков — гистогенез, органогенез, системогенез. Из зародышевых листков образуется:
I. ЭКТОДЕРМА:
1)эпидермис кожи и его производные (сальные, потовые, молочные железы, ногти, волосы), нервная ткань, нейросенсорные и сенцоэпителиальные клетки органов чувств, эпителий ротовой полости и его производные (слюнные железы, эмаль зуба, эпителий аденогипофиза), эпителий и железы анального отдела прямой кишки;
II. МЕЗОДЕРМА:
1) дерматомы — собственно кожа (дерма кожи);
2) миотомы — скелетная мускулатура;
3) склеротомы — осевой скелет (кости, хрящи);
4) нефрогонотомы (сегментные ножки) — эпителий мочеполовой системы;
5) спланхнотомы — эпителий серозных покровов (плевра, брюшина, околосердечная сумка), гонады, миокард, корковая часть надпочечников;
6) нефрогенная ткань — эпителий нефронов почек.
III. ЭНТОДЕРМА:
1) часть энтодермы, образованная из прехордальной пластинки — эпителий и железы пищевода и дыхательной системы;
2) часть энтодермы, образованная из гипобласта — эпителий и железы всей пищеварительной трубки (включая печень и поджелудочную железу); участвует при образовании переходного эпителия мочевого пузыря (аллантоис).
IV. МЕЗЕНХИМА:
1) все виды соединительной ткани (кровь и лимфа, рыхлая и плотная волокнистая соед.ткань, соед.ткань со специальными свойствами, костные и хрящевые ткани);
2) гладкая мышечная ткань;
3) эндокард.
ПЛАЦЕНТА
При формировании плаценты участвуют со стороны плода трофобласт и внезародышевая мезенхима. А со стороны матери — функциональный слой слизистой матки. Трофобласт и внезародышевая мезенхима образуют хорион. Это происходит следующим образом: вначале трфобласт представляет собой полый пузырек из одного слоя клеток, в последующем клетки трофобласта начинают усиленно размножаться и поэтому трофобласт становится многослойным. Причем клетки наружных слоев сливаются друг с другом и образуют симпласт — этот слой называется симпластическим трофобластом; самый внутренний слой трофобласта сохраняет клеточное строение и называется клеточным трофобластом (цитотрофобласт). Параллельно с этим из эмбриобласта выселяются клетки — внезародышевая мезенхима и она покрывает внутреннюю поверхность цитотрофобласта. Эти 3 слоя вместе (симпластический и клеточный трофобласт, внезародышевая мезенхима) назваются хорионом или сосудистой оболочкой.
В дальнейшем симпластический трофобласт по всему периметру хориона образует выросты — I ворсинки хориона; I ворсинки хориона начинают выделят протеолитические ферменты, которые разрушают эпителий матки и через образовавшуюся бреш зародыш внедряется в толщу слизистой матки, т.е. происходит имплантация; эпителий матки за зародышем восстанавливается и поэтому зародыш оказывается замурованным в толще слизистой матки.
Все 3 слоя хориона вместе образуют II ворсинки хориона, которые проникают через стенки кровеносных сосудов слизистой матки и плавают в крови матери, т.е. начинается плацентация. В дальнейшем во II ворсинки хориона врастают сосуды плода и II ворсинки превращаются в III ворсинки. Кровь плода в сосудах плода в III ворсинках и кровь матери не смешиваются, между ними находится плацентарный барьер, который состоит из следующих слоев:
1. Эндотелий капилляров плода в III ворсинках.
2. Базальная мембрана капилляров плода.
3. Внезародышевая мезенхима.
4. Цитотрофобласт.
5. Симпластический трофобласт.
Типы плацент у млекопитающих:
1. Эпителиохориальная — ворсинки хориона проникают в просвет маточных желез, эпителий не разрушается (пример: у свиньи).
2. Десмохориальная — ворсинки хориона проникают через эпителий матки и контактируют с рыхлой соед.тканью эндометрия (пример: у жвачных).
3. Эндотелиохориальная — ворсинки хориона проникают через эпителий матки и прорастают в стенку сосудов матери до эндотелия, но в просвет сосуда не проникают (пример: у хищников).
4. Гемахориальная — ворсинки хориона проходят через эпителий матки, прорастают через стенки сосудов матери и плавают в крови матери, т.е. ворсинки контактируют непосредственно с кровью матери (пр.: человек).
ПРОВИЗОРНЫЕ ОРГАНЫ — это временные органы, функционируют только в эмбриональном периоде. К ним относятся: хорион, амнион, желточный мешок, аллантоис и серозная оболочка.
1. Хорион — строение и функции смотри выше.
2. Амнион — образуется из внезародышевой эктодермы и мезенхимы (у птиц еще и париетальный листок спланхнотомов). Функция — создает благоприятную защитную водную среду вокруг зародыша.
3. Желточный мешок — образуется из внезародышевой энтодермы и мезенхимы (у птиц еще и висцеральный листок спланхнотомов). Функции: обеспечивает питание зародыша; там образуются первые кровеносные сосуды, первые клетки крови и половые клетки — гонобласты.
4. Аллантоис («мочевой мешок») — это слепое выпячивание энтодермы в заднем отделе первичной кишки; в нем накопливается шлаки обмена плода, т.е. выделительная функция; у млекопитающих является проводником пупочных сосудов плода и участвует при формировании эпителия мочевого пузыря.
5. Серозная оболочка — имеется только у птиц, образуется из внезародышевой эктодермы и париетального листка спланхнотомов; основная функция — обеспечение дыхания зародыша, кроме того выполняет защитную функцию.
У млекопитающих, и в том числе у человека, хорошо выражены и активно функционируют хорион и амнион, а желточный мешок и аллантоис плохо выражены (рудементарны); серозная оболочка у млекопитающих отсутствует.
|
|
|
|
Дата добавления: 2014-01-20; Просмотров: 577; Нарушение авторских прав?; Мы поможем в написании вашей работы!