КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Введение. Жизнь на Земле представляет собой форму существования материи
|
|
|
|
Жизнь на Земле представляет собой форму существования материи. Живая материя зародилась спонтанно, то есть самопроизвольно, как закономерный результат космических процессов и явилась завершением химической эволюции – естественного образования и накопления органических соединений. Жизнь можно определить как активное поддержание и самовоспроизведение специфической структуры материи, идущее с затратой полученной извне энергии. Из этого определения вытекает необходимость постоянной связи организмов с окружающей средой, осуществляемой путем обмена веществом и энергией. Современная наука не располагает прямыми доказательствами того, как и где возникла жизнь. Существуют лишь косвенные свидетельства, полученные путем экспериментов, и данные из области палеонтологии, геологии, палеоклиматологии, астрономии, биохимии. Наиболее известны два основных взгляда на место и характер зарождения жизни. Суть первого сводится к абиогенному (то есть вне организма) возникновению живого в условиях формирующейся Земли. Теорию такого рода в 20-х годах ХХ столетия выдвинули А.И.Опарин и Дж.Холдейн. Этим взглядам наиболее соответствует мнение о том, что жизнь на Земле монофилетична, то есть ведет начало от единого предка.
Согласно другим гипотезам местом возникновения жизни считается Космос, откуда зачатки живого могли быть занесены на Землю с веществом метеоритов, комет или иным образом (метеоритная бомбардировка Земли закончилась около 4 млрд. лет назад). Такого рода гипотезы тесно связаны с идеей полифилетического, то есть неоднократного, зарождения жизни и в свое время поддерживались создателем учения о биосфере В.И.Вернадским.
Возможность абиогенного синтеза органических соединений типа аминокислот, пуринов, пиримидинов, сахаров в условиях восстановительной атмосферы древнейшей Земли в 50-60-х годах ХХ в. была подтверждена экспериментально, но в то же время сложные органические молекулы найдены в околозвездном пространстве и могли быть занесены на Землю из Космоса.
Однако сложность решения вопроса связана не с доказательствами возможностей синтеза органики на Земле или в Космосе, а с проблемой возникновения генетического кода. Важный и до сего времени нерешенный вопрос состоит в том, каким образом органические молекулы организовались в системы, способные к самовоспроизведению.
Живое вещество характеризуется некоторыми типичными чертами. Главнейший признак живого – дискретность, то есть существование в виде отдельных организмов (особь, индивидуум). Каждый организм представляет собой открытую целостную систему, через которую, как явствует из определения жизни, проходят потоки вещества и энергии. Поэтому нередко говорят не просто о живом веществе, но о живых системах.
Неотъемлемое свойство любой живой системы – обмен веществ, или метаболизм. Параллельно метаболизму в любом организме осуществляются постоянное превращение энергии и ее обмен.
Для живых организмов характерно самовоспроизведение, обеспечивающее непрерывность и преемственность жизни.
Живые организмы – самоорганизующиеся и саморегулируемые системы. Благодаря саморегуляции устанавливаются на определенном уровне различные физиологические процессы. Организмы являются открытыми термодинамическими системами, способными к любому обмену веществом и энергией. Без поступления энергии извне эти системы не могут существовать и поддерживать свою целостность.
Перечисленные основные свойства определяют сложность живых систем, а также способность самостоятельно поддерживать и увеличивать относительно высокую степень упорядоченности в среде с меньшей упорядоченностью.
Основу живого вещества составляют два класса химических соединений – белки и нуклеиновые кислоты. Белки ответственны за обмен веществ и энергии в живой системе, то есть за все реакции синтеза и распада, протекающие в организме непрерывно. Нуклеиновые кислоты обеспечивают хранение и передачу наследственной информации, то есть способность живых систем к самовоспроизведению. Они являются матрицей, содержащей полный набор информации, на основе которого синтезируются видоспецифические белки клетки.
В состав живых организмов также входят липиды (жиры), углеводы. Органические вещества других классов встречаются у представителей отдельных групп организмов.
В живых системах найдены многие химические элементы, присутствующие в окружающей среде. Однако для жизни необходимо около 20 из них. Эти элементы получили название биогенных, поскольку постоянно входят в состав организмов и обеспечивают их жизнедеятельность. В среднем около 70% сырой массы организмов составляет кислород (O), 18% – углерод (C), 10% – водород (H). Далее следуют азот (N), кальций (Ca), калий (K), фосфор (P), магний (Mg), сера (S), хлор (Cl), натрий (Na). Это универсальные биогенные элементы, присутствующие в клетках всех организмов и называемые макроэлементами. Часть элементов содержится в организмах в крайне низких концентрациях (до тысячных долей процента), но они также необходимы для нормальной жизнедеятельности (микроэлементы). Их функции и роль очень разнообразны. Многие микроэлементы входят в состав ферментов, некоторые влияют на рост. Насчитывается до 30 микроэлементов – металлов (Al, Fe, Cu, Mn, Zn, Mo, Co, Ni, Sr) и неметаллов (I, Se, Br, F, As, B).
Присутствие в клетках биогенных элементов зависит от особенностей организма, от состава среды, пищи, экологических условий, в частности от растворимости и концентрации солей в почвенном растворе. Недостаточность или избыточность биогенных элементов приводит к ненормальному развитию организма или даже к его гибели. Добавки биогенных элементов в почву для создания их оптимальных концентраций широко используются в сельском хозяйстве.
Обмен веществ, или метаболизм, – это совокупность протекающих в организмах химических превращений, обеспечивающих их рост, развитие, жизнедеятельность, воспроизведение, постоянный контакт и обмен с окружающей средой. В ходе обмена веществ происходит расщепление и синтез молекул, входящих в состав клеток, образование, разрушение и обновление клеточных структур и межклеточного вещества.
Обмен веществ сводится к двум противоположным, но одновременно взаимосвязанным процессам: анаболизму и катаболизму. Первый сводится к построению веществ тела в результате реакций синтеза с потреблением энергии. Второй объединяет реакции распада с высвобождением энергии. Процессы синтеза и распада белков, нуклеиновых кислот, липидов, углеводов и аскорбиновой кислоты получили название первичного обмена, или первичного метаболизма. Они свойственны всем живым существам и играют решающую роль в поддержании их жизнедеятельности. Образование и превращение прочих классов органических соединений относятся к вторичному метаболизму. Вторичный метаболизм наиболее обычен для растений, грибов и ряда прокариот (от греческого «про» – перед, «карион» – ядро), то есть организмов, не имеющих морфологически оформленного ядра. Процессы вторичного метаболизма и сами вторичные метаболиты часто играют существенную адаптивную (приспособительную) роль у организмов, лишенных способности к перемещению в пространстве.
Организмы поддерживают свое существование и целостность, получая энергию извне. Накапливается эта энергия в виде энергии химических связей. Наиболее энергоемкими являются жиры, углеводы, менее энергоемкими – белки. Универсальный источник энергии для всего живого на Земле – энергия солнечной радиации, но способы использования ее живыми организмами различны. Зависят от световой энергии фотоавтотрофные организмы (зеленые растения и фототрофные прокариоты). Они запасают энергию, образуя первичные органические соединения из неорганических в процессе фотосинтеза. Гетеротрофные организмы (животные, грибы, большинство прокариот) не могут создавать органические соединения из неорганических. В качестве источника углерода они используют органические формы этого элемента. В качестве источника энергии они также используют органические вещества, созданные в процессе жизнедеятельности фотоавтотрофами. Х емоавтотрофные организмы (некоторые прокариоты) получают энергию, выделяемую при перестройке молекул минеральных или органических соединений в процессе химических реакций. Источником углерода для разных групп хемоавтотрофов служат также минеральные формы углерода.
Высвобождение энергии осуществляется в процессе распада органических соединений чаще всего с помощью двух процессов – брожения и дыхания.
Индивидуальное развитие отдельного организма от зарождения до смерти получило название онтогенеза. Отдельные онтогенезы в цепи поколений складываются в единый последовательный процесс, называемый гологенезом. Совокупность онтогенезов, то есть гологенез, лежит в основе эволюции. Под эволюцией подразумевается процесс необратимого исторического развития живой природы и отдельных его звеньев, ведущий к усложнению или упрощению организации живого. В эволюционном процессе различают микроэволюцию и макроэволюцию.
Под микроэволюцией подразумевают процессы видообразования, сопровождающиеся изменением генетического состава популяций, формированием адаптаций к меняющейся среде.
Макроэволюция – это образование таксонов выше ранга вида. Ход макроэволюции определяется микроэволюцией. Макроэволюция реализуется в филогенезе, то есть в процессе исторического становления и развития отдельных видов и других систематических групп более высокого ранга. Как и вся эволюция, филогенез связан с онтогенезом и гологенезом. Этот процесс принято изображать графически в виде филогенетического древа (или филемы), показывающего возможные родственные связи между отдельными ветвями живого (или филами). Ход филогенеза чаще всего подчиняется определенным правилам, называемым правилами эволюции (рис. 1.1).
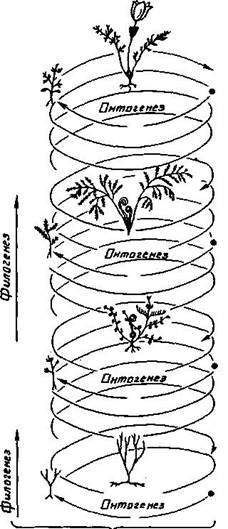
Рис. 1.1. Схема соотношения онтогенеза и филогенеза (пояснения терминов в тексте).
На Земле существует около 2-2,5 млн. видов организмов и около 500 млн. видов вымерло в предшествующие геологические эпохи. Однако при таком многообразии живого можно выделить несколько разных уровней строения и изучения живой материи. Главнейшие уровни строения живого: молекулярно-генетический, онтогенетический, популяционно-видовой и биогеоценотический. На каждом уровне строения живая материя характеризуется специфическими элементарными структурами и элементарными явлениями.
На молекулярно-генетическом уровне гены представляют элементарные структуры, а элементарными явлениями можно считать их способность к конвариантной редупликации – самовоспроизведению с изменениями на основе матричного принципа и к мутациям.
На онтогенетическом уровне элементарной структурой живого следует считать особь, индивид, а элементарным явлением – онтогенез, или развитие особи от зарождения до смерти.
Основу популяционно-видового уровня представляет популяция, а процесс свободного скрещивания (панмиксия) – элементарное явление.
Биогеоценотический уровень жизни характеризуется элементарной структурой – биогеоценозом, а обмен веществ и энергии в биогеоценозе составляет элементарное явление.
При изучении живой материи выделяют несколько уровней ее организации:
1. молекулярный;
2. клеточный;
3. тканевой;
4. органный;
5. онтогенетический;
6. популяционный;
7. видовой;
8. биогеоценотический;
9. биосферный.
Каждому уровню соответствует особая биологическая наука, несколько биологических наук или раздел биологии.
На молекулярно-генетическом уровне живые организмы исследуются молекулярной биологией и генетикой; на клеточном – цитологией; на тканевом и органном – анатомией и морфологией, а также физиологией; на онтогенетическом – морфологией и физиологией; на популяционном – популя-ционной генетикой; на видовом – систематикой и эволюционным учением; на биогеоценотическом – геоботаникой, экологией, биогеоценологией; на биосферном – биогеоценологией.
Земля сформировалась как плотное тело около 4,6 млрд. лет назад. Этой цифрой датируется начало так называемого гадейского эона (надэры). Нет геологических доказательств, подтверждающих существование жизни на Земле в это время, но несомненно, что живое возникло или было занесено на Землю именно в гадее, поскольку в архейских отложениях в начале следующего эона уже встречаются разнообразные организмы. Предполагается, что обогащение водоемов в конце гадея аминокислотами, пуриновыми и пиримидиновыми основаниями, сахарами создало так называемый «первичный бульон», служивший источником питания древнейшим гетеротрофам.
Архейский эон (надэра), или архей, охватывает период времени от 3900 до 2600 млн. лет назад. К этому времени относится возникновение древнейших осадочных пород, образованных частицами, осаждавшимися из водной среды, часть которых сохранилась в районе рек Лимпопо (Африка), Исуа (Гренландия), Варавууна (Австралия), Алдана (Азия). Эти породы содержат биогенный углерод, связанный в своем происхождении с жизнедеятельностью организмов, а также строматолиты и микрофоссилии. Строматолиты – кораллоподобные осадочные образования (карбонатные, реже кремниевые), представляющие собой продукты жизнедеятельности древнейших автотрофов. В протерозое они всегда связаны с цианобактериями, но их происхождение в архее не вполне ясно. Микрофоссилии – микроскопические включения в осадочные породы ископаемых микроорганизмов.
В архее все организмы относились к прокариотам. Часть из них, очевидно, была гетеротрофами-деструкторами (разрушителями), использовавшими органические вещества, растворенные в «первичном бульоне» и превращавшими их в процессе жизнедеятельности в простые соединения типа Н2О, СО2 и NН3. Другая часть микроорганизмов архея составила группу продуцентов – организмов, способных к осуществлению либо аноксигенного фотосинтеза (фотосинтеза без выделения кислорода), либо хемосинтеза.
На стадии аноксигенного фотосинтеза остались современные пурпурные и зеленые серные фотобактерии. Донором электронов в процессе фотосинтеза у них служил главным образом Н2S, а не Н2О. Микроорганизмы-продуценты могли уже фиксировать атмосферный азот.
Получение энергии у большинства архейских организмов осуществлялось путем брожения или специфического анаэробного дыхания, при котором источником кислорода, отсутствующего в атмосфере, служили сульфаты, нитриты, нитраты и другие соединения.
Древнейшие бактериальные биоценозы – сообщества живых организмов, включавшие только продуцентов и деструкторов, были похожи на пленки плесени (так называемые бактериальные маты), располагавшиеся на дне водоемов или в их прибрежной зоне. Оазисами жизни часто служили вулканические районы, где на поверхность из литосферы поступали водород, сера и сероводород – основные доноры электронов. Геохимический цикл (круговорот веществ), существовавший на планете до возникновения жизни и наиболее ярко проявлявшийся, очевидно, в циркуляции атмосферы, пополнился биогеохимическим циклом. Биогеохимические циклы (круговорот веществ, связанный с организмами), совершавшиеся при помощи продуцентов – аноксигенных фотосинтетиков и деструкторов, были относительно простыми и осуществлялись главным образом в форме восстановленных соединений типа сероводорода, аммиака.
В протерозойском эоне, или протерозое, который начался 2600 млн. и закончился 570 млн. лет назад, такое положение изменилось. Ископаемые остатки и разнообразные следы жизни в осадочных породах этого времени довольно обычны. Строматолиты образуют мощные многометровые толщи и их существование в протерозое связывают с жизнедеятельностью цианобактерий. Эта новая группа продуцентов появилась на арене жизни в самом начале протерозойского эона или даже в конце архея. Она обладала способностью к оксигенному фотосинтезу, то есть могла использовать Н2О в качестве донора электронов, при этом свободный кислород выделялся в атмосферу. Появление цианобактерий привело к преобразованиям всей биосферы Земли. Восстановительная атмосфера Земли превратилась в окислительную. Анаэробное живое население планеты постепенно сменилось на аэробное. Концентрация кислорода в результате жизнедеятельности цианобактерий постепенно повышалась и примерно 2 млрд. лет назад достигла 1% от современной. Атмосфера стала окислительной. Это послужило предпосылкой развития аэробного хемосинтеза и эволюционно самого молодого из процессов получения энергии – аэробного дыхания. Существенно изменяются и усложняются биогеохимические циклы. Накопление кислорода стало препятствием для циркуляции элементов в форме восстановленных соединений. Бактериальные архейские сообщества строгих анаэробов заменяются цианобактериальными сообществами (цианобактериальные маты), в которых главенствующую роль играют фотосинтезирующие прокариоты.
Изменение характера атмосферы оказалось главной предпосылкой появления строгих аэробов эукариот – этого важнейшего биологического события середины протерозоя. Первые эукариоты появились около 1,8 млрд. лет назад и были, по-видимому, планктонными, или свободноплавающими организмами. Древние эукариотические организмы могли быть как гетеротрофами, так и автотрофами, пополнявшими две основные, ранее существовавшие экологические группы продуцентов и деструкторов. Длительное время в протерозое прокариоты и эукариоты существовали совместно в составе альгобактериальных сообществ (сообществ, где компонентами были эукариотические водоросли и бактерии), заменивших 1,4 млрд. лет назад цианобактериальные сообщества.
Происхождение эукариот объясняют различно. Традиционная точка зрения связывает их появление с постепенным усложнением структуры прокариотической клетки. Согласно другой теории, которая разделяется сейчас большинством биологов, эукариоты возникли как итог внутриклеточного симбиоза древних безоболочечных анаэробных микроорганизмов с разными типами оксифотобактерий. Проблема возникновения эукариотных организмов до конца не решена. Существуют различные гипотезы происхождения эукариотической организации живой материи. Одной из таких гипотез является гипотеза эндосимбиоза (симбиогенеза). Эта теория была выдвинута в конце XIX – начале XX в. Современный этап ее развития связан с работами американского биолога Линн Маргелис, которая предполагает, что эукариотическая клетка возникла в результате нескольких последовательных эндосимбиозов (симбиотического существования одной клетки внутри другой) древних безоболочечных анаэробных прокариот, способных к процессу брожения, с различными прокариотическими аэробами. Эукариотные клетки сформировались в результате симбиоза между чрезвычайно далекими друг от друга видами прокариот: нуклеоцитоплазма образовалась из организмов-«хозяев», митохондрии – из бактерий, дышащих кислородом, пластиды эукариот. На первом этапе эндосимбиоза возникли различные одноклеточные эукариотические простейшие, которые в процессе эволюции дали начало многоклеточным эукариотам из царств грибов, растений и животных. Общая схема процесса эндосимбиоза показана на рис. 1.2.
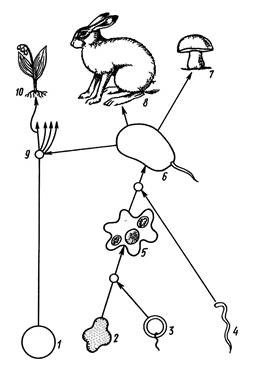
Рис. 1.2. Схема происхождения эукариотических клеток путем эндосимбиоза (по Л.Маргелис, с изменениями): 1 – разные группы оксифотобактерий, обладающие различными пигментами (предшественники хлоропластов), 2 – термоплазмы (термостойкие прокариоты), 3 – подвижные нефотосинтезирующие прокариоты (предшественники митохондрий), 4 – подвижные спирохеты или спироплазмы (предшественники жгутиков), 5 – гетеротрофная амебоидная эукариотическая клетка, 6 – древнейшая эукариотическая подвижная клетка, обладающая жгутиком, 7 – царство грибов, 8 – царство животных, 9 – зона нескольких предполагаемых симбиозов подвижной эукариотической клетки с различными группами оксифотобактерий; возникли различные линии эволюции растений, одна из них дала начало высшим растениям (10).
В конце протерозоя, очевидно, существовали многоклеточные растения и грибы, но их ископаемые остатки не сохранились. Древнейшие многоклеточные организмы появились примерно 950 млн. лет назад. С этого времени начинают исчезать строматолиты, а экологические системы Земли стали сложнее на одно звено. В них, помимо продуцентов и деструкторов, включились консументы – потребители органического вещества живых организмов. Еще до начала четвертого эона – фанерозоя, уже существовали сообщества, в которых преобладали планктонные (свободноплавающие) и бентосные (донные) водоросли и многоклеточные растительноядные животные. Роль цианобактерий и других прокариот в формировании основной массы биогеоценозов позднего протерозоя была незначительной.
Фанерозойский эон, или фанерозой (надэра явной жизни), начался примерно 570 млн. лет назад и продолжается до настоящего времени. Осадочные толщи фанерозоя изобилуют ископаемыми животными и растениями. Само начало фанерозоя датируется по появлению в ископаемых остатках большого числа многоклеточных животных, имеющих внутренние или наружные скелеты. Фанерозой принято делить на три эры: палеозойскую, или эру древней жизни, мезозойскую – эру средней жизни и кайнозойскую – эру новой жизни.
Особенность истории развития живых организмов в фанерозое состояла в том, что определенным группам животных соответствовали определенные группы растений. Это и понятно, поскольку основу для развития животных создавало процветание тех или иных растительных сообществ. Поэтому эволюция растений шла с некоторым опережением эволюции животных.
Древнейшие наземные растения риниофиты появились в конце силура (410-420 млн. лет назад). Во второй половине девона – карбоне (430 – 300 млн. лет назад) возникли все основные группы (таксоны) ныне живущих и вымерших растений, кроме покрытосеменных (цветковых). Однако господствующими формами в течение всего палеозоя, начиная с середины девона, были споровые: хвощевидные, плауновидные и папоротниковидные, древовидные формы которых нередко образовывали леса. Голосеменные появились на Земле не позднее верхнего карбона (290 млн. лет назад), но их господство начинается с конца перми (около 220 млн. лет назад) и продолжается в течение почти всего мезозоя до середины мела. В нижнем мелу, примерно 145-120 млн. лет тому назад, появляются покрытосеменные, которые к середине верхнего мела занимают господствующее положение. Это положение они сохранили в течение всего кайнозоя до нашего времени (рис. 1.3).
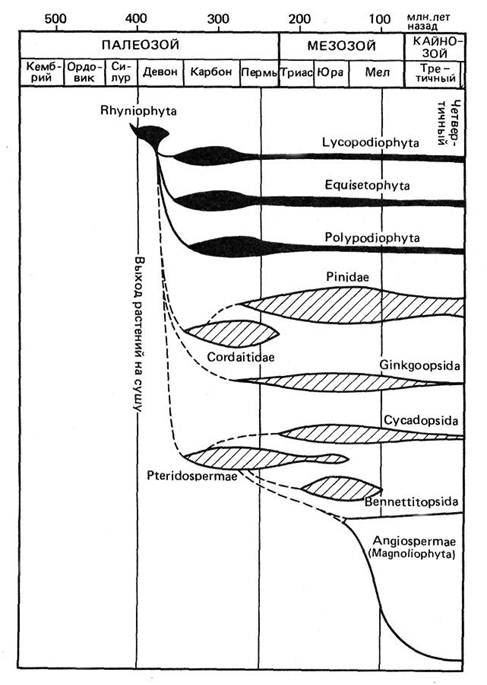
Рис. 1.3. Эволюционный возраст основных филогенетических групп растительного мира.
Со времени К.Линнея (XVIII в.) в науке господствовала система двух основных групп организмов (или царств органического мира): растений (Vegetabilia, или Р1апtае) и животных (Аnimalia). Однако открытие в XX в. ряда важных различий в метаболизме и ультраструктуре клетки у разных групп организмов побудило биологов изменить устоявшийся взгляд. Начиная с середины 50-х годов ХХ в. широко обсуждаются другие возможные системы (Р.Уиттейкер, Г.Кёртис, Ч.Джефри, Е.Додсон, А.Тахтаджян, Я.Старобогатов). Количество выделяемых царств в этих системах колеблется от трех до десяти. В основу деления живого на царства положены способы питания, особенности ультраструктуры митохондрий и пластид, химический состав клеточных оболочек и основных запасных веществ клеток, некоторые другие принципы.
Ниже приведен краткий перечень крупнейших систематических трупп, позволяющий представить значимость и положение в общей системе живого изучаемых в курсе “Ботаника” таксонов.
Империя неклеточные организмы ( Noncellulata ). Представители не имеют морфологически оформленной клетки. Империя включает одно царство вирусы (Virae).
Империя клеточные организмы ( Сellulata ). Представителиимеют морфологически оформленную клетку. Включает две подимперии.
1. Подимперия доядерные ( Procaryota ) – не имеют морфологически оформленного ядра. Объединяет два царства:
а) Царство архебактерии (Archaebacteria) – в основе клеточных оболочек имеют кислые полисахариды без муреина;
б) Царство настоящие бактерии, или эубактерии (Eubacteria) – в качестве основного структурного компонента клеточных оболочек содержится гликопротеид муреин.
2. Подимперия ядерные или эукариоты ( Eucaryota ) – имеют морфологически оформленное ядро. Подразделяется на четыре царства:
а) Царство протоктисты (Protoctista) – автотрофы или гетеротрофы; тело не расчленено на вегетативные органы; отсутствует стадия зародыша; гаплоидные или диплоидные организмы; включает водоросли и грибоподобные организмы.
б) Царство животные (Animalia) – гетеротрофы; питание путем заглатывания или всасывания; отсутствует плотная клеточная стенка; диплоидные организмы; имеется чередование ядерных фаз.
в) Царство грибы (Fungi, Mycota) – гетеротрофы; питание путем всасывания; имеется плотная клеточная стенка, в основе которой хитин; гаплоидные или дикарионтические организмы; тело не расчленено на органы и ткани;
г) Царство растения (Plantae) – автотрофы; питание за счет процесса аэробного фотосинтеза; имеется плотная клеточная оболочка, в основе которой целлюлоза; характерно чередование полового (гаметофит) и бесполого поколения (спорофит) с преобладанием диплоидного поколения. К растениям относятся ископаемые риниофиты и зостерофиллофиты, а также современные моховидные, хвощевидные, плауновидные, папоротниковидные, голосеменные и покрытосеменные.
Объектом изучения ботаники являются в первую очередь представители царства растения, фототрофные протоктисты – водоросли. В то же время в данном курсе будут рассматриваться отдельные вопросы по морфологии и систематике некоторых групп фотоавтотрофных прокариот (цианобактерии), а также грибов и грибоподобных организмов. Эти систематические группы еще недавно считались представителями царства растения.
У многих растений и водорослей функции полового и бесполого размножения выполняют разные поколения, которые нередко представлены морфологически различными особями. Соотношения этих двух поколений у основных групп показаны на рис. 1.4.
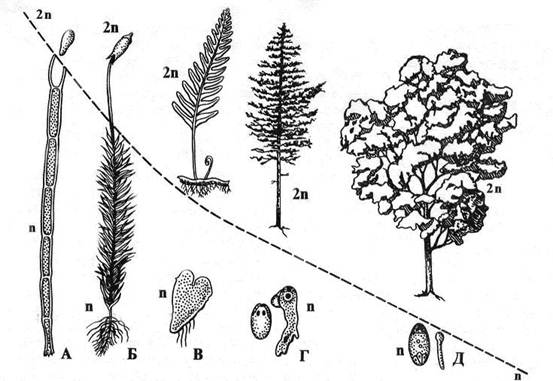
Рис. 1.4. Соотношение и строение полового и бесполого поколений в жизненных циклах. А – водоросли; Б – мхи, В – папоротники, Г – голосеменные, Д – покрытосеменные (цветковые).
Каждая из основных групп организмов является предметом изучения самостоятельной биологической науки или комплекса близких наук. В частности, бактерии (исключая цианобактерии, которые традиционно исследовались ботаниками-альгологами, то есть специалистами по водорослям) изучаются бактериологией или наукой более широкого плана – микробиологией, предметом интереса которой служат все микроскопические живые организмы. Протистология исследует простейших, то есть одноклеточных, колониальных и многоклеточных эукариот, имеющих дотканевую организацию. Микология (от греческого «микес» – гриб) изучает представителей царства грибов. Ботаника изучает царство растений и автотрофных прокариот. Наконец, зоология занимается животными организмами. Особое царство составляют доклеточные формы жизни – вирусы (Virae). Наука о вирусах называется вирусологией.
Ботаника (от греческого «ботанэ» – растение, трава) – комплекс биологических наук о растениях. Первые датируемые сведения о растениях содержатся в клинописных таблицах Древнего Востока. Основы ботаники как науки заложили древние греки. Древнегреческий философ и естествоиспытатель Теофраст (около 370-285 лет до н. э.) назван К. Линнеем «отцом ботаники». После общего упадка естествознания в средние века ботаника начинает интенсивно развиваться с XVI в.
В XVIII – XIX вв. происходит развитие и дифференциация ботаники на отдельные ботанические дисциплины и к первой половине XX в. складывается весь комплекс наук о растениях. Основной раздел ботаники – систематика растений. Систематика описывает все ископаемые и современные растительные организмы, разрабатывает классификацию и создает научную основу для изучения филогении растений, то есть выявляет родство таксонов.
Морфология исследует особенности и закономерности внешнего строения растений. Основные успехи в этой области знаний были достигнуты преимущественно в XIX и XX вв. Исследование внутренней структуры растений – задача анатомии, которая зародилась в середине XVII в. после изобретения микроскопа, но, подобно морфологии, главнейшие открытия были также сделаны в XIX и XX вв.
Эмбриология – ботаническая дисциплина, изучающая закономерности образования и развития зародыша растений. Основы эмбриологии заложены во второй половине XVIII в., но фундаментальные открытия были сделаны к началу XX в.
Физиология тесно связана с морфологией и биохимией растений. Начало физиологии было положено опытами по питанию растений, осуществленными во второй половине XVIII в. Ныне это активно развивающаяся наука, занимающаяся изучением происходящих в растениях процессов: фотосинтеза, транспорта веществ, водного обмена, роста, развития, дыхания.
География растений зародилась в начале XIX в. Она занимается изучением основных закономерностей пространственного распространения таксонов (видов, родов и более высоких) и растительных сообществ на Земле. Из ботанической географии к концу XIX в. выделилась геоботаника – наука, исследующая основные закономерности формирования, состава, структуры и функционирования растительных сообществ, а также особенности их пространственного распределения.
Экология растений выясняет отношение растительных организмов к факторам среды и взаимоотношения растений с другими организмами. Она возникла на стыке экологии и ботаники на рубеже XIX и XX вв. и в настоящее время это одна из важнейших отраслей знаний о природе.
Помимо фундаментальных ботанических дисциплин, выделяют ряд прикладных наук, также относимых к ботанике. Главнейшей из них считается ботаническое ресурсоведение, или экономическая ботаника. Она рассматривает все аспекты использования растений человеком.
В зависимости от объектов и методов их изучения, а также практических потребностей выделяют ряд других ботанических дисциплин. В пределах морфологии растений выделяют карпологию – раздел знаний о плодах, из анатомии – палинологию, изучающую пыльцу и споры. Предметом исследования палеоботаники являются ископаемые растения. У палеоботаники свои методы изучения, близкие к методам палеонтологии.
Альгология изучает водоросли, бриология – мхи, птеридология – папоротники.
Особая роль растений в жизни на Земле состоит в том, что без них было бы невозможно существование животных и человека. Зеленые растения являются основной группой организмов, способных аккумулировать энергию Солнца, создавая органические вещества из неорганических. При этом растения извлекают из атмосферы диоксид углерода (углекислый газ) и выделяют кислород, поддерживая ее постоянный состав. Будучи первичными продуцентами органических соединений, растения являются определяющим звеном в сложных цепях питания большинства гетеротрофов, населяющих Землю.
Благодаря фотосинтезу и непрерывно действующей трансформации биогенных элементов создается устойчивость всей биосферы Земли и обеспечивается ее нормальное функционирование.
Обитая в различных условиях, растения образуют растительные сообщества (фитоценозы), обусловливая разнообразие ландшафтов и экологических условий для других организмов. При участии растений формируются почва, торф; скопления ископаемых растений образовали бурый и каменный уголь. Глубокие нарушения растительности неизбежно влекут за собой необратимые изменения биосферы и отдельных ее частей и могут оказаться гибельными для человека как биологического вида.
Существует пять основных сфер, где прямо или косвенно используются растения:
1) в качестве продуктов питания для человека и корма для животных,
2) как источники сырья для промышленности и хозяйственной деятельности,
3) как лекарственные средства и сырье для получения медицинских препаратов,
4) в декоративном озеленении,
5) в охране и улучшении окружающей среды.
Пищевое значение растений общеизвестно. В качестве продуктов питания человека и корма для животных, как правило, используются части, содержащие запасные питательные вещества или сами вещества, извлеченные тем или иным образом. Потребность в углеводах в основном удовлетворяется за счет крахмал- и сахаросодержащих растений. Роль источников растительного белка в рационе человека и животных выполняют в основном некоторые растения из семейства бобовых. Плоды и семена многих видов используют для получения растительных масел. Большинство витаминов и микроэлементов также поступает вместе со свежей растительной пищей. Существенную роль в питании людей играют пряности и растения, содержащие кофеин – чай и кофе.
Техническое использование растений и продуктов из них осуществляется по нескольким основным направлениям. Наиболее широко применяются древесина и волокнистые части растений. Ценность древесины определяется потребностью в ней при изготовлении деревянных конструкций любых типов и при производстве бумаги. Сухая перегонка древесины позволяет получить значительное количество важных органических веществ, широко употребляемых в промышленности и в быту. Во многих странах древесина – один из основных видов топлива. Остро стоит вопрос о замене угля и нефти энергетически богатыми веществами, продуцируемыми некоторыми растениями.
Несмотря на широкое распространение синтетических волокон, растительные волокна, получаемые из хлопчатника, льна, конопли, джута, липы сохранили большое значение при производстве многих тканей. Для лечебных целей растения применяют очень давно. В народных и традиционных медицинах они составляют основную массу лекарственных средств. В научной медицине России примерно треть препаратов, применяемых для лечения, получают из растений. Считается, что с лечебными целями народы мира используют не менее 21000 видов растений и грибов. В России около 55 видов лекарственных растений культивируется. Подробно с использованием растений в медицине студенты ознакомятся в курсах фармакогнозии и фармакологии. Не менее 1000 видов растений разводят в декоративных целях.
Функционирование всех экологических систем биосферы, частью которой является и человек, целиком определяется растениями. Растительные ресурсы относятся к категории восполняемых (при правильной эксплуатации) в противоположность, например, невосполняемым минеральным ресурсам. Чаще всего растительные ресурсы делят на ресурсы природной флоры (сюда относятся все дикие виды) и ресурсы культивируемых растений. По объему и значимости в жизни человечества они существенно различаются. Природные ресурсы флоры ограничены и в их естественном объеме могли бы обеспечить питанием лишь около 10 млн. человек. Наиболее широко дикорастущие растения используются в качестве технических источников сырья, в хозяйственной деятельности человека, а также как источник лекарственных средств. Появление культурных растений и возникновение дополнительных растительных ресурсов связано со становлением древнейших человеческих цивилизаций. Существование этих цивилизаций могло обеспечиваться только определенным «ассортиментом» окультуренных растений, дающих необходимое количество растительных белков, жиров и углеводов. Жизнь современного человека и современная цивилизация невозможны без широкого использования культивируемых растений. Почти все культурные растения (примерно 1500 видов) относятся к покрытосеменным. К середине XX в. культивируемые растения занимали 15 млн. км2, то есть около 10% всей поверхности суши Земли.
Наращивание ресурсов культурных растений возможно в весьма широких пределах как за счет увеличения площадей их возделывания (экстенсификации), так и за счет улучшения агротехники и выведения высокопродуктивных сортов (интенсификация). Считается, что полная мобилизация восполняемых ресурсов, включая растительные, может обеспечить существование на Земле не менее 6 млрд. человек.
Переходящие к земледелию народы часто независимо друг от друга вводили в культуру растения окружающей их дикой флоры. Можно выделить ряд основных центров древнейшего земледелия, называемых еще центрами происхождения культурных растений. Учение о центрах происхождения культурных растений впервые было разработано Н.И.Вавиловым (1887-1943). Согласно его представлениям существовало восемь таких центров. В настоящее время выделяют десять центров происхождения культурных растений (рис. 1.5).
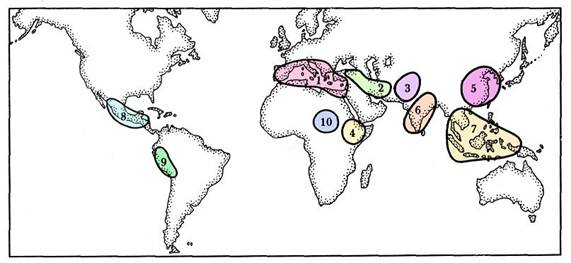
Рис. 1.5. Центры происхождения культурных растений (по Н.И.Вавилову, с изменениями): 1 – средиземноморский, 2 – переднеазиатский, 3 – среднеазиатский, 4 – эфиопский, 5 – китайский, б – индийский, 7 – индонезийский, 8 – мексиканский, 9 – перуанский, 10 – западносуданский.
У европеоидных народов с примыкающей к ним группой эфиопов отмечены четыре центра: средиземноморский, переднеазиатский, эфиопский, среднеазиатский. Монголоиды имели один центр – северокитайский. У австралоидных народов юго-востока и юга Азии земледелие развилось автохтонно (то есть независимо) в двух очагах: индийском и индонезийском (или индомалайском). У американских народов возникли мексиканский и перуанский центры. Негроидные народы тропической Африки имели один основной центр земледелия – западносуданский.
Средиземноморский центр объединяет области Европы, Африки и Азии, прилегающие к Средиземному морю. Это родина некоторых сортов овса, льна, мака, белой горчицы, маслины, рожкового дерева, капусты, моркови, свеклы, лука, чеснока, спаржи, редьки.
Переднеазиатский очаг расположен в Малой Азии, Закавказье, Иране. Это родина пшеницы однозернянки и двузернянки, твердой пшеницы, ржи, ячменя.
Среднеазиатский (Центральноазиатский) центр охватывает бассейны Сырдарьи и Амударьи, индийское Пятиречье (формирующее реку Инд). Он является родиной мягкой пшеницы, гороха, чечевицы, нута, маша, возможно, конопли, сарептской горчицы, винограда, груши, абрикоса и яблони.
Эфиопский центр – Эфиопия и Сомали. Это родина сорго, кунжута, клещевины, кофейного дерева, некоторых форм овса, финиковой пальмы.
Китайский центр располагается в области умеренного пояса бассейна реки Хуанхэ. Здесь сформировались культуры проса, гречихи, сои, ряда листопадных плодовых деревьев, таких, как хурма, китайские сорта сливы и вишни.
Индийский центр находится на полуострове Индостан. Основными культурами древнего земледелия этого очага были тропические виды, часть которых затем продвинулась в страны умеренного климата. Индийский центр – родина риса, азиатских хлопчатников, манго, культурных форм огурца и баклажана.
Индонезийский центр занимает главным образом территорию современной Индонезии. Здесь была родина ямса, хлебного дерева, мангустана, бананов, дуриана и, возможно, кокосовой пальмы. Тропическое садоводство получило отсюда своих важнейших представителей. В Индонезийском очаге взяты в культуру такие важные пряные растения, как черный перец, кардамон, гвоздичное дерево, мускатный орех.
Мексиканский центр включает большую часть территории Центральной Америки. Отсюда человечество получило маис (кукурузу), обыкновенную фасоль, красный стручковый перец, хлопчатники Нового света (так называемые упленды), махорку и, вероятно, папайю, или дынное дерево.
Перуанский (Южноамериканский) центр занимает территорию Перу, Эквадора, Боливии, Чили, отчасти Бразилии. Из этого очага в культуру взяты картофель, томат, длинноволокнистый «египетский» хлопчатник, ананас и табак. В новейшее время отсюда вывезено и окультурено хинное дерево.
Западносуданский центр расположен на части территории тропической Африки. Отсюда началась культура масличной пальмы, орехов кола, ряда тропических зернобобовых.
Около 30% всех выпускаемых медицинских препаратов готовят из лекарственного растительного сырья. Источником сырья служат как дикорастущие, так и культивируемые растения. Это определяет целый комплекс проблем, в которых провизор обязан квалифицированно разобраться. Прежде всего, он должен уметь узнавать и характеризовать растения, что делает строго необходимым хорошее знание их морфологии и систематики. Подлинность лекарственного растительного сырья в процессе фармакогностического анализа определяется на основе изучения различных макроскопических и микроскопических признаков. Обязательным разделом всех стандартов, регулирующих качество лекарственного сырья, является подробная макроскопическая и микроскопическая характеристики. Макроскопический анализ предполагает хорошее знание морфологии растений и владение соответствующей ботанической терминологией. При микроскопическом анализе провизоры-аналитики изучают растительное сырье анатомически. В этом случае им помогает знание анатомии растений. К анатомическим исследованиям объектов нередко прибегают при судебно-медицинских экспертизах в тех случаях, когда на месте преступления обнаруживаются растительные остатки.
Изучение физиологии растений позволяет понять суть процессов, которые приводят к образованию в растениях продуктов первичного и вторичного обмена (метаболизма). Многие из них оказываются фармакологически активными и используются в медицинской практике. С культивированием лекарственных растений провизор сталкивается относительно редко, но заготовки дикорастущего лекарственного растительного сырья осуществляются многими аптеками. Поэтому знание флоры региона необходимо для правильного планирования и организации заготовок. В последние десятилетия различные причины привели к истощению главнейших естественных ресурсов ряда лекарственных растений в традиционных районах заготовок. Актуальными сделались ресурсные исследования по выявлению новых промышленных массивов лекарственных растений и инвентаризация запасов лекарственного растительного сырья. Эти работы осуществляют провизоры-фармакогносты. Выполнение ресурсных исследований невозможно без знания местной флоры, элементов ботанической географии и владения основными геоботаническими методами. Наконец, провизор обязан выполнять главнейшие природоохранные мероприятия, которые должны учитываться при сборе растительного сырья. Это залог длительной эксплуатации зарослей дикорастущих лекарственных растений.
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 757; Нарушение авторских прав?; Мы поможем в написании вашей работы!