КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Лекция №5 Гормональная система растений
|
|
|
|
Растительная клетка как часть многоклеточного организма, должна не только выполнять свои функции, направленные на поддержание собственной целостности, но и функционировать согласованно с другими клетками. Именно поэтому между клетками должен происходить обмен информацией. Одна группа клеток при этом становится "отправителями" сигнала, а другая группа клеток воспринимает его. Если сигнал имеет химическую природу, молекулу, выполняющую сигнальную функцию, называют первичным мессенджером (от англ. " message "-"послание","сообщение").
Растительная клетка воспринимает разнообразные химические стимулы и реагирует на них. Так, изменение концентрации сахаров во флоэмном токе, олгигомерные сахара и фрагменты хитина, индолилуксусная кислота, брассиностероиды и др. вызывают физиологическую реакцию. Среди этого довольно обширного списка химических веществ можно выделить гормоны растений.
Для того, чтобы вещество отнести к гормонам, необходимо, чтобы оно обладало следующими свойствами.
1. Вещество вызывает специфический физиологический ответ у определенных клеток. Особенность растительных гормонов в том, что они запускают крупные программы развития не только на уровне клеток, но и на уровне тканей, органов, целого растения.
2. Вещество синтезируется в самом растении одной группой клеток, а отвечает на него другая (разобщено место синтеза и место действия, т.е. необходим транспорт сигнального вещества по растению). Как правило, фитогормоны являются низкомолекулярными соединениями (не более 2кДа). Возможно, это связано с необходимостью транспорта по растению. Отметим, что к синтезу фитогормонов потенциально способна любая клетка растений.
3. Вещество практически не играет роли в основном метаболизме клетки, используется только для сигнальных целей.
Кроме того, иногда эти свойства расширяют, добавляя дополнительные качества.
4. Вещество должно действовать в низкой концентрации (низкой считают концентрацию начиная с 10-5моль/л).
Все эти правила введены для того, чтобы ограничить круг веществ, которые традиционно считают растительными гормонами. Однако, проще перечислить вещества, которые в научной литературе принято называть гормонами. Основных групп классических гормонов пять.
1. Ауксины. 2. Цитокинины. 3. Гиббереллины. 4. Абсцизины.
5. Этилен.
Часто к этому списку добавляют и другие вещества: брассиностероиды, жасминовую кислоту, салициловую кислоту, некоторые фенольные соединения и др.
На самом деле всеми классическими свойствами обладают немногие гормоны (например, ауксины). Абсцизины обычно действуют в точке синтеза, распространяясь лишь на небольшое расстояние. Этилен транспортируется только в виде предшественника; фенолы, салициловая и жасминовая кислоты действуют в концентрациях, превышающих 10-5 моль/л.
Кроме гормонов, к сигнальным молекулам можно отнести олигосахарины (так называют олигосахариды с физиологической активностью), лектины (белки, способные вступать во взаимодействие с полисахаридами) и короткие пептиды. Действующая концентрация этих веществ порой очень низка - порядка 10-12═ - 10-15 моль/л.
Ауксины - гормоны роста побега.
История открытия ауксинов.
В 1880 году Чарльз Дарвин и его сын Френсис Дарвин поставили перед собой проблему: каким органом растения воспринимают свет. Растения, стоящие на подоконнике, поворачиваются к солнцу, побеги и листья изгибаются в сторону наибольшего освещения. Причем изгиб происходит в зоне чуть пониже верхушки. Дарвины с помошью светонепроницаемой ширмы изолировали от света субапикальную область, в которой обычно происходит изгиб. При боковом освещении они обнаружили, что растения изгибаются к свету, хотя зона изгиба не доступна солнечным лучам. Незакрытой оставалась только верхушка (стеблевой апекс) проростка, и логично было ее накрыть черным колпачком. Действительно, после этого изгиба в сторону света не наблюдается, хотя зона изгиба освещена солнцем с одной стороны. Результаты Дарвинов неоспоримо говорили о том, что направление света воспринимает верхушка проростка (апекс) и передает информацию о направлении света в нижележащую зону. Дарвины предположили, что верхушка вырабатывает химическое соединение, которое транспортируется вниз и вызывает неравномерный рост. Гипотетическое вещество Дарвины назвали ауксином (от греч. ауксо - расту).
Гипотеза Дарвинов была не единственным объяснением наблюдавшихся явлений. Так, сигнал мог передаваться в виде электрических импульсов и приводить к тому же результату. Выдвинутое Дарвинами предположение нуждалось в эксперимнтальной проверке.
Через 50 лет после опытов Дарвинов, наш соотечественник Холодный и его немецкий коллега Вент независимо поставили новую серию экспериментов, чтобы изучить природу сигнала, передающего информацию из апекса побега в субапикальную зону изгиба.
Вещество, передающее сигнал из одной части растения в другую, должно быть хорошо растворимым в воде. Значит, если отрезать много побеговых апексов и поместить их в воду, то в раствор вместе с другими веществами перейдет и гипотетический ауксин. Затем нужно сделать так, чтобы с одной стороны в проростке оказалось больше "ауксина", а с другой - меньше. Для этого верхушку с собственными ауксинами нужно удалить, а на срез с какого-нибудь края нанести водный экстракт.
Однако, работать с водным раствором было трудно - экстракт растекается по срезу. Тогда опыт видоизменили. Апексы располагали на поверхности тонкого слоя агарового геля. У подопытного растения удаляли верхушку и сбоку накладывали на срез кубик агара, в котором предположительно содержался ауксин. Растения всегда изгибались в сторону, противоположную наложенному агаровому блоку. В качестве контроля использовали агаровый блок, не пропитанный экстрактом побеговых апексов. В этом случае изгиба не происходило.
Холодный и Вент сделали вывод, что гипотетический ауксин действительно существует и вызывает растяжение клеток. Можно было предположить, что при одностороннем освещении ауксин перетекает на затененную сторону и вызывает изгиб к свету.
Не имея в руках выделенного препарата чистого ауксина, Вент и Холодный изучили основные свойства этот сигнального вещества. Так, если вырезать зону изгиба и накладывать агаровый кубик снизу (с базальной стороны), то изгиба не будет. Это означает, что ауксин движется по растению полярно (т.е. в строго заданном направлении) - от апекса побега к его основанию, а затем - к кончику корня. Была измерена скорость транспорта ауксинов. Для этого на участок заданной длины помещали сверху агар с ауксином, а с низу - агаровый блок без ауксина. Верхний агаровый блок служил донором ауксинов, а нижний - своеобразным коллектором. Нижний блок через равные промежутки времени заменяли новым. Всю полученную серию нижних блоков анализировали на проростках на предмет наличия ауксинов.Скорость транспорта ауксинов обычно составляет от нескольких миллиметров до нескольких сантиметров в час.
Однако, необходимо было выделить ауксин в чистом виде. Немецкий химик Кёгль в 1939 году исследовал химический состав мочи вегетарианцев. Он отметил, что в отличие от мочи других людей, она содержит новое вещество, которое удалось выделить и расшифровать его формулу. Это была индолилуксусная кислота (ИУК). Чтобы научиться определять ее в малых количествах и изучить ее физиологические свойства, Кёгль испробовал ИУК в разных живых системах. Выяснилось, что ИУК способна усиливать рост проростков с отрезанной верхушкой, как и ауксин. Если наложить агаровый блок с ИУК асимметрично, то происходит характерный изгиб субапикальной зоны.
Лишь через 6 лет из верхушек побегов было выделено то же самое соединение - индолилуксусная кислота - и стало ясно, что она и является одним из акусинов. К настоящему времени вопос о других соединениях ауксиновой природы окончательно не решен. Известен ряд веществ, обладающих ауксиновой активностью (фенилуксусная кислота, индолилпировиноградная кислота), но их активность ниже, чем у ИУК и они менее распространены.
Кроме них химикам удалось синтезировать вещества, вызывающие такой же физиологический эффект, как и природные ауксины. Поскольку они не встречаются в растениях, их называют синтетическими аналогами ауксино в. Среди них упомянем лишь 2,4-Дихлорфеноксиуксусную кислоту (2,4-Д) и a-нафтилуксусную кислоту (НУК). Синтетические аналоги эффективно связываются с рецепторами ауксина, однако слабо взаимодействуют с системой транспорта или окислительной деградации естественных ауксинов.
Биосинтез и деградация ауксинов.
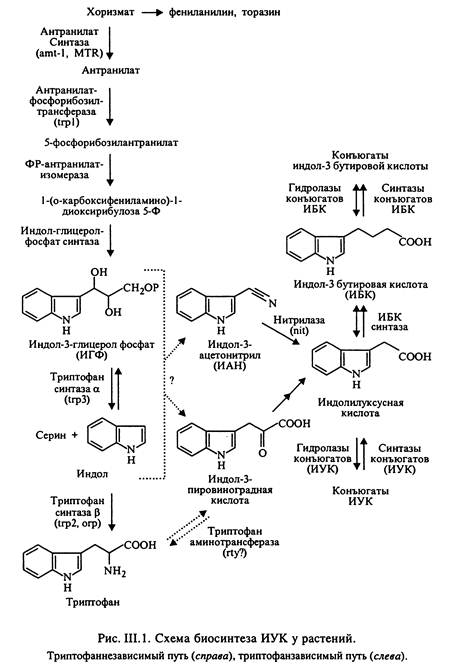
Ауксины синтезируются у разных растений разными путями. Долгое время считалось, что ИУК образуется единствыенным путем из аминокислоты триптофана. Этот процесс происходит как минимум в три стадии: (1) декарбоксилирования, т.е. удаления СО2; (2) дезаминирования, т.е. удаления NH2-группы; (3) окисления, т.е. удаления электроно. Некоторые растения не ограничиваются этим набором реакций, а включают в метаболизм ауксинов соединения серы, и биосинтез растягивается на 6 и более стадий.
Есть еще один путь синтеза ИУК, в котором триптофан не участвует. Некоторые растения способны эксплуатировать оба пути: триптофановый и "нетриптофановый" и переключают их в зависимости от обстоятельств (см рис.).
После образования ауксин часто подвергается дальнейшим превращениям. Так, ИУК может связываться с сахарами, аминокислотами или даже белками, образуя не активные (запасные) формы. При необходимости ИУК можно освободить из этих соединений и восстановить ее физиологическую активность. Интересно, что в классических опытах Дарвинов с канареечной травой синтеза ауксина в верхушке колеоптиля не происходит ИУК образуется в период роста зародыша семени, а когда зерновки созревают, ИУК переходит в форм неактивного гликозида. При прорастании гликозиды ИУК транспортируются в апекс проростка, где происходит высвобождение активного ауксина, и уже свободная ИУК распределяется по колеоптилю канареечной травы в зависимости от освещенности.
Кроме образования неактивных конъюгатов, ИУК может разрушаться. Это происходит либо специфически (с помощью ИУК-оксидазы), либо неспецифически (полифенолоксидазой, которая кроме ИУК может работать с разнообразными фенольными соединениями). В обоих случаях потеря ауксина невосполнима.
Таким образом, по мере удаления от точки синтеза концентрация ауксинов падает за счет необратимого окисления и связывания в неактивные формы.
Мутанты.
Для изучения генетического контроля метаболизма и функции ауксинов используют ауксиновые мутанты. Выделяют два типа мутантов:
1. Мутанты с нарушением синтеза. Такие мутанты называют гормонзависимые или гормондефицитные. Характерной чертой мутантов этой группы является восстановление нормального фенотипа при добавлении гормона, т.е. имеется возможность получать фенокопии дикого типа. 2. Мутанты с нарушением чувствительности к гормону. Среди них выделяют устойчивые (resistant) и чувствительные (sensitive) формы: устойчивые характеризуются способностью роста на максимальных концентрациях гормона, в то время как чувствительные формы не способны расти на более низких концентрациях гормона по сравнению с диким типом, при этом ткани или целые растения погибают.
Самок большое количество ауксиновых мутантов получено у арабидопсиса, табака и томатов. Ауксиновые мутанты, как правило, меньше по размеру, с маленькими сморщенными листьями, укороченным черешком, замедленной геотропической реакцией, плохо развитой корневой системой; с повышенной кустистостью и недоразвитием сосудистой системы.
Ауксинустойчивые мутанты: AXR-1-4 (auxin-resistens), TIR-1-3 (transport inhibitor response), MP (monopteros), SUR (superroot), Agr-1-3 (нарушение гравитропизма) и др.
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 2096; Нарушение авторских прав?; Мы поможем в написании вашей работы!