КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Компоненти мононуклеотидів
|
|
|
|
| Тип нуклеїнових кислот | Пентоза | Пуринові основи | Піримідинові основи |
| РНК ДНК | Рибоза Дезоксирибоза | Аденін, гуанін Аденін, гуанін | Цитозин, урацил Цитозин, тимін |
Схематичне позначення:

| – азотиста основа |
| – 5- вуглецевий цукор (пентоза) |

| – фосфорний залишок |
| – нуклеозид |
| – мононуклеотид |
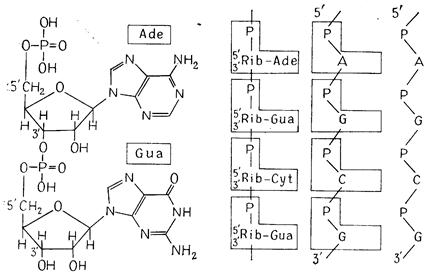
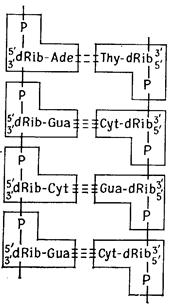
Гідроксильна група (ОН) в атома 3'-С пентози одного мононуклеотида може етерифікуватись фосфатом іншого нуклеотида, і в результаті утвориться динуклеотид (рис. 2.5). У такий спосіб приблизно від 70 до >108 мононуклеотидів зв'язуються в полінуклеотидний ланцюг. Осьовий кістяк цієї молекули складається із залишків, що чергуються, фосфату й пентози, тоді як основи приєднані збоку.
Всі пентозні залишки одного ланцюга орієнтовані атомом 5'-С у одному напрямку, а атомом 3'-С - у протилежному. Таким чином, кожний ланцюг полярний: він має 5'-кінець (фосфатний кінець) і 3'-кінець (гідроксильний кінець) (рис. 2.6, 2.7).
| ||
| А | Б | В |
Рис. 2.6. Структура РНК. А. Динуклеотид з аденозин- і гуанізинмонофосфат. Б, В, Відрізок молекули РНК із чотирма мононуклеотидними залишками; зліва направо - усе більш спрощені схеми.
2.3.2 ДНК
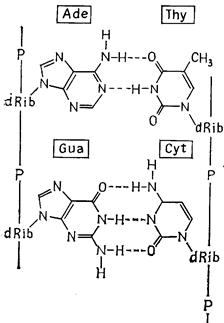
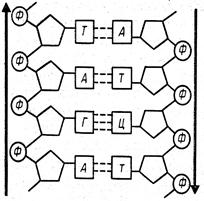
Молекули ДНК складаються приблизно з 2000-108 і більше мононуклеотидів. У кожній молекулі два полінуклеотидні ланцюги об'єднані в один подвійний ланцюг. Основи розташовані парами друг проти друга й з’єднані водневими зв'язками. По стереохімічним причинам до утворення пар здатний тільки комплементарні ("ті, що підходять один до одного") основи: аденін і тимін з'єднуються між собою двома, а гуанін і цитозин - трьома водневими зв'язками (рис. 2.7, 2.8). Два ланцюги розташовуються антипаралельно: 5'-кінець одного ланцюга лежить проти 3'-кінця іншого. Ланцюги комплементарні по азотистих основах: наприклад, послідовності G-А-C-G-Т-G протистоїть в іншому ланцюзі послідовність C-T-G-C-А-C.
Молекула ДНК містить інформативні й неінформативні ділянки. В інформативних ділянках послідовність основ (первинна структура) являє собою матеріальний еквівалент генетичної інформації. Кожне повідомлення закодоване специфічною послідовністю з чотирьох знаків - A, G, С і Т, подібно тому, як письмові повідомлення кодуються знаками (буквами) алфавіту або абетки Морзе.
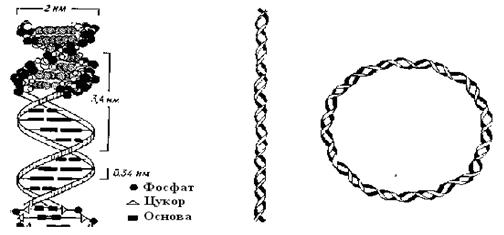
Вторинна структура ДНК - це подвійна спіраль: два полінуклеотидних ланцюги закручені навколо спільної уявлюваної осі, і між ними утворюються дві спіральні борозенки неоднакової глибини (рис. 2.7, 2.8). Подвійна спіраль стабілізована водневими зв'язками й гідрофобними взаємодіями. Її діаметр 2 нм,
| 
| 
|
| А | Б | В |
Рис. 2.7. Структура ДНК. А. Відрізок подвійної спіралі ДНК із двома мононуклеотидними залишками в кожному з ланцюгів. Б. Відрізок подвійної спіралі ДНК із чотирма мононуклеотидними залишками в кожному ланцюзі.
В. Схема подвійної спіралі ДНК із 4-ма мононуклеотидними залишками
крок спирали 3,4 нм; кожен виток містить 10 пар нуклеотидів, так що кожна пара займає 0,34 нм по осі спіралі (рис. 2.8).
| ||
| А | Б | В |
Рис. 2.8. Конформація ДНК. А. Схема подвійної спіралі. Б. Спрощене зображення подвійної спіралі. В. Кільцева подвійна спіраль
У хромосомах подвійні спіралі утворюють тяжі із двома кінцями. Подвійні спіралі ДНК в цитоплазматичних органоїдах і у без'ядерних (прокаріотичних) організмів, замкнуті в кільце (рис. 2.8), У хромосомах подвійні спіралі з'єднані з білками (в основному іонними зв'язками) і утворюють разом з ними третинну структуру, а також структури більше високого порядку.
Підвищення температури (приблизно до 90°С) або зміна рН приводить до денатурації ДНК. Полінуклеотидні ланцюги роз'єднуються і в граничному випадку переходять у повністю неупорядкований стан, так званий "статистичний клубок".
2.3.3 РНК і пре- РНК
Молекули РНК, як правило, одноланцюгові (рис. 2.9). Однак у більшості випадків при петле подібному складанні молекул окремі комплементарні одна одній ділянки одного й того ж ланцюга в результаті спарювання основ утворюють спіралі (рис. 2.9); при цьому А з'єднується з U, a G із С.
Існують різні типи РНК, що розрізняються по величині молекул, структурі й функції (табл. 2.2). Всі види РНК (за винятком вірусних) синтезуються на молекулі ДНК як копії ділянок цієї молекули (транскрипція). При цьому спочатку утворюється більш довгий попередник - первинний транскрипт, що потім перетворюється в більше коротку РНК. Це перетворення включає ряд етапів і називається процесінгом. Первинний транскрипт і проміжні продукти процесінга відомі за назвою пре-РНК.
При процесінгу, по-перше, ланцюг РНК коротшає в результаті відщепления відрізків від його кінців або ж видалення якоїсь ділянки із середини ланцюга з наступним з'єднанням частин, що залишилися (сплайсинг); по-друге, приєднуються нові кінцеві послідовності нуклеотидів; по-третє, відбувається модифікація нуклеотидів, наприклад їх метилювання або гідрування.
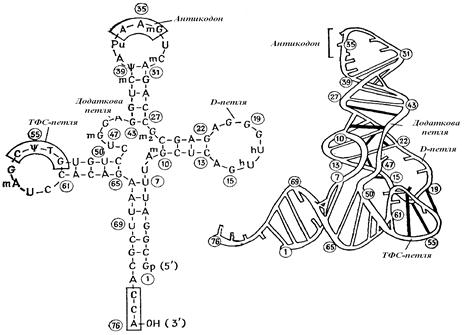
Рис. 2.9. Структура tPHK. Фенілаланінова t PHK: первинна й вторинна структура (ліворуч) і третинна структура (праворуч).
Приблизно 10% всієї клітинної РНК становлять відносно низькомолекулярні t PHK (табл. 2.2). Ці РНК не пов'язані з якими-небудь частками. При реалізації генетичної інформації кожна t PHK приєднує й переносить певну амінокислоту.
Таблиця 2.2
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 722; Нарушение авторских прав?; Мы поможем в написании вашей работы!