КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Температура
|
|
|
|
Продолжительность развития организмов зависит от температуры, которая в природных условиях изменяется в широких пределах. Для учета температурного фактора при определении продукции и дыхания гидробионтов используется «температурный коэффициент» Вант-Гоффа (Q10). Цифровое значение которого непосредственно указывает. Во сколько раз ускоряются жизненные процессы при увеличении температуры на 100.
at = 1 / Q10(20-T)/10
где температурный множитель; Т – температура воды в период наблюдений; 20 – значение температуры воды, для которых выведено абсолютное большинство расчетных формул. Значения Q10 находятся по «нормальной кривой» Крога для следующих диапазонов температуры:
| Т0С | Q10 |
| 5-10 | 3,5 |
| 10-15 | 2,9 |
| 15-20 | 2,5 |
| 20-25 | 2,3 |
Динамика роста популяций одноклеточных водорослей.
Скорость роста клеток зависит от скорости биохимических реакций, обуславливающих синтез белков и нуклеиновых кислот, которые в отличие от запасных веществ являются активной частью протоплазмы. Суммарным выражением скорости биосинтетических реакций популяции одноклеточных водорослей может служить скорость деления клеток, т.к. рост отдельной клетки ограничен известным пределом. Рост водорослей рассмотрим на примере фитопланктона, т.к. он является основным продуцентом ОВ в водных экосистемах.
В условиях культур у всех видов водорослей различают два возраста – индивидуальный и популяционный, т.к. скорость деления клеток тесно связана с происхождением индивидуального и популяционного циклов (пояснить на примере диатомовых).
Популяционный возраст описывается, как правило, S-образной кривой. Во время роста культуры водорослей в лимитированных объемах различают следующие фазы роста:
1. Лаг-фаза, в которой не происходит увеличения клеток;
2. Экспоненциальная, или логарифмическая, фаза, в которой численность клеток увеличивается в геометрической прогрессии;
3. Фаза уменьшения относительного роста;
4. Стационарная фаза, когда количество клеток постоянно;
5. Фаза гибели.
Все фазы являются результатами взаимодействия клетки со средой.
Между концентрацией веществ снаружи и внутри клеток существует определенная зависимость, что дает возможность ускорять или замедлять определенные энзиматические процессы ускорять или замедлять определенные ферментативные процессы внутри клетки, тем самым изменяя определенным образом концентрацию продуктов обмена в окружающей среде.
Продолжительность лаг-фазы зависит от популяционного возраста инокулята (увеличиваясь с его возрастом), а также от концентрации в нем клеток.
В лаг-фазе происходит реставрация энзимов и увеличение концентраций внеклеточных органических метаболитов в среде, продуцируемых самими клетками до оптимального уровня, при котором начинается быстрый рост. В частности, рост водорослей стимулирует гликолиевая кислота.
После окончания лаг-фазы водоросли переходят в экспоненциальную фазу, которая характеризуется быстрым их ростом. Продолжительность этой фазы зависит в основном от скорости деления и величины максимальной концентрации клеток, при которой способен расти данный вид (у большинства диатомовых водорослей эта фаза заканчивается при достижении концентрации 10-15 тыс. кл/мл). Чем медленнее размножаются водоросли, тем более продолжительной становится экспоненциальная фаза роста.
Третья – пятая фазы роста наступают вследствие полного поглощения биогенных элементов в процессе роста и происходит накопление в среде аутотоксичных веществ. Торможение роста водорослей в культурах в большей мере вызывают продукты жизнедеятельности самих водорослей.
* * * * *
ЗАДАЧА:
Определить суточную валовую продукцию фитопланктона в единицах энергии, если:
Vсклянки = 250 мл
Продолжительность экспозиции = 3 часа
Продолжительность светлого времени суток = 14 часов
∆О2 («светлых» склянок) = 0,45 мл
∆О2 («темных» склянок) = 0,21 мл
1 мл О2 = 20,34 Дж
РЕШЕНИЕ:
1) Чистая продукция в «светлое» время: 0,45 мл: 3 часа ∙ 14 часов = 2,1 мл
2) Дыхание ночью: 0,21 мл: 3 часа ∙ 10 часов = 0,7 мл
3) Суточная валовая продукция в склянке: (2,1 мл + 0,7 мл) ∙ 20,34 Дж = 56,95 Дж/сут
4) Суточная валовая продукция: 56,95 Дж/сут ∙ 1000/250 = 227,8 Дж∙л-1∙сут-1
ЗАДАЧА:
Определить суточную чистую продукцию фитопланктона в единицах энергии, если:
Vсклянки = 250 мл
Продолжительность экспозиции = 1 сутки
Продолжительность светлого времени суток = 14 часов
∆О2 («светлых» склянок) = 0,30 мл
∆О2 («темных» склянок) = 0,15 мл
1 мл О2 = 20,34 Дж
РЕШЕНИЕ:
1) Чистая суточная продукция в склянке = 0,30 мл ∙ 20,34 Дж = 6,10 Дж/сут
2) Чистая суточная продукция: 6,10 Дж/сут ∙ 1000/250 = 24,4 Дж∙л-1∙сут-1
* * * *
Вопросы:
- Первичная продукция и методы ее определения.
- Валовая и чистая продукция.
- Основные факторы, определяющие первичную продукцию.
- Динамика роста популяций одноклеточных водорослей.
Лекция 3. Вторичная продукция
Общие закономерности роста животных. Методы изучения роста и возраста животных. Размножение и плодовитость. Способы определения продукции популяций и биоценозов.
Основные понятия и показатели
Продуктивность вторичная – общее количество ОВ, произведенного совокупностью всех гетеротрофных организмов (бактерий, грибов, животных) на единицу площади за единицу времени. Как и первичную продукцию ее делят на валовую и чистую. Однако. Применительно ко вторичной продукции нет оснований различать валовую и чистую продукции. Поскольку по определению, вторичная продукция представляет собой именно чистую продукцию.
Удельная продукция – продукция за единицу времени в пересчете на единицу биомассы продуцирующей биосистемы.
Впервые Р/В коэффициент использовал Р. Демоль в 1927 году, имея в виду отношение продукции животных к биомассе пищевых организмов. Коэффициент Р/В как отношение продукции к биомассе той же системы ввел Л.А. Зенкевич (1931) в период, когда были возможны лишь грубо ориентировочные оценки только годовой продукции крупных сообществ и групп, таких как, как планктон, бентос, рыбы. Обозначение показателя отражало единственный возможный тогда способ его вычисления: продукцию оценивали за год, а затем делили на биомассу, получая годовую Р/В. Оперируя большими промежутками времени, многие исследователи стали относить продукцию то к средней, то к начальной, то к минимальной биомассе за год. Показатели оказывались несравнимыми, возникала путаница.
В настоящее время рекомендуется вычислять Р/В для возможно более кратких отрезков времени (например, сутки) и при этом относить продукцию к средней биомассе.
Детализация исследований позволила установить, что суточная величина Р/В популяции представляет собой средневзвешенную величину суточных удельных весовых приростов всех особей популяции. Следовательно, о величине Р/В популяции можно судить по данным о скорости роста особей и возрастной структуре популяции. Таким образом, принципиально меняется рекомендуемая последовательность исследования продукционных показателей.
Если раньше использовали путь: биомасса – расчет продукции за год – вычисление годового Р/В, то теперь ясно, что гораздо эффективнее иной путь: биомасса – расчет суточного Р/В по данным о скорости роста и возрастной структуре – определение продукции как произведение биомассы на удельную продукцию. Знания о продуктивности системы при таком подходе оказываются гораздо более полными.
Разумеется, и сейчас во многих случаях суточную удельную продукцию приходится вычислять делением годовой величины на 365 дней. Но дело не только в способе вычисления этого показателя. При математическом моделировании оперировать с выражением Р/В неудобно. Чтобы убедиться в этом, достаточно записать равенство
Р = В ∙ Р/В
При такой форме записи оно выглядит как простое тождество, ничего не дающее исследователю. Однако его широко используют для расчета продукции. Следовательно, мы возвращаемся к факту, что Р/В не обязательно вычислять делением продукции на биомассу, отношение может быть получено независимым путем по эколого-физиологическим данным и затем использовано для расчета продукции.
С учетом сказанного для рассматриваемого показателя используется термин «удельная продукция», обозначая ее СW.
Вместе с тем, для выведения обобщающей ("универсальной") зависимости, позволившей описать функционирование беспозвоночных разных видов на различных этапах жизненного цикла, была использована безразмерная величина удельной интенсивности дыхания, подобно удельной продукции, широко применяемой в гидробиологии. Эта величина выражается отношением скорости поглощения кислорода к массе животного. В частности, данный показатель можно получить, если потребление кислорода и массу тела выразить в одинаковых единицах, например в энергетических (джоулях).
Методы изучения роста и возраста животных.
При изучении роста водных организмов используют обычно следующие методы: 1) наблюдения в лабораторных условиях или в садках, выставляемых в местах обитания животных; 2) наблюдения в естественных условиях по изменениям роста меченных животных; 3) определение приростов по фиксированным возрастным признакам (морфологическим, структурным, физико-химическим); 4) анализ размарно-возрастной структуры популяций.
1. Наблюдения в лабораторных условиях. Проводят периодические промеры и взвешивания животных, на основании которых судят о характере и скорости их роста. При лабораторных экспериментах следует строго соблюдать неизменные условия внешней среды, особенно температуру и обеспеченность пищей, а также ее качество.
Время наблюдений может быть сокращено за счет одновременного использования в экспериментах животных, относящихся к разным возрастным группам. Данные такого рода не дают представления о росте конкретной особи, но характеризуют осредненную зависимость линейных размеров или массы от времени роста некоторой средней особи данной группы.
2. Наблюдения в естественных условиях. Наблюдения за ростом меченных животных в природных условиях проводят на основе периодических отловов животных с их измерением. Полученные данные обычно осложняются малым возвратом животных при последующих поимках. Этот метод можно использовать применительно к крупным животным с прочными наружными покровами или наружным скелетом (например, крупные моллюски, ракообразные, рыбы). В качестве меток используются различные красители, краски, бирки и т.п.
3. Определение приростов по фиксированным признакам. Основаны на анализе циклических элементов, чаще всего так называемых колец или линий роста, которые отмечаются, например, на раковинах моллюсков, чешуе или отолитах рыб. Эти элементы образуются через определенные периоды времени – обычно через год. Эти методы требуют точного определения периодичности формирования указанных образований и установления продолжительности временного цикла.
4. Анализ гистограмм частотно-размерного состава. Достоверные результаты с помощью данного метода могут быть получены в том случае, если на гистограммах каждый возрастной класс может быть уверенно выделен в любые периоды времени. Наиболее четко различимы на гистограммах размерно-возрастные группы в популяциях быстро растущих животных, для которых характерен сжатый период нереста. В силу этого на гистограммах таких видов молодь составляет единую размерную группу, надежно отличающуюся по размерам от животных предыдущего поколения. У животных, медленно растущих, с большой продолжительностью жизни и растянутым по времени нерестом чаще всего размерные группы трудно различимы. На гистограммах таких видов обычно наблюдается множество зубцов, трудно соотносимых с возрастом.
Данный метод наиболее часто используется для планктонных ракообразных. В частности М.Б. Ивановым был разработан метод определения продолжительности развития особи планктонных копепод по относительному количеству в планктоне животных на отдельных стадиях развития. Численность рачков каждой стадии рассчитывается в процентах от суммарной численности всей популяции. Время между максимумами численности двух смежных стадий равно полусумме продолжительности развития каждой из них. Необходимо установить продолжительность прохождения первой науплиальной стадии с тем, чтобы по данным проб, собранных в водоеме, определить продолжительность развития особей всех остальных стадий.
Общие закономерности роста животных.
Продукционный процесс в популяции можно интерпретировать как прирост биомассы, включающий рост массы особей и увеличение их числа за счет размножения. Исходя из основного балансового равенства, продукцию в одних случаях можно выразить через соотношение усвоенной пищи и трат на обмен, в других – через динамику численности или рост особей.
Оценка продукции на основе анализа роста массы имеет ведущее значение. Это объясняется тем, что изучение роста массы и возрастной структуры популяций требует меньшей затраты сил и времени, дает обычно более надежные исходные данные для продукционных расчетов, чем сопряженное со многими трудностями составление «таблиц жизни» и оценка на их основе скорости увеличения популяции. Большинство вариантов расчета продукции животных осуществляется по данным о росте.
Типы роста животных
Общие принципы изучения роста животных, в том числе и гидробионтов, были подробно разработаны Г.Г. Винбергом. Для водных животных различают три типа роста: экспоненциальный, параболический и сигмоидальный, или S–образный, каждый из которых определенным образом связан со скоростью энергетического обмена у животных. Названия типов роста определяются видом функций и кривых, с помощью которых могут быть описаны эмпирические данные о росте животных.
Представление об основных типах роста разработаны на основе анализа кривых роста. Знание кривой роста позволяет оценить суточные абсолютные и удельные приросты особей различного возраста (массы). Эти данные используются при расчетах продукции популяций.
Основу удельной продукции популяции составляют удельные скорости роста особей. Среднюю удельную скорость роста массы Cw за период t2 – t1 можно рассчитать по формуле:
Cw = 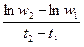 =
= 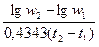 [1]
[1]
где w1 и w2 – масса в момент t1 и t2 соответственно. Это уравнение справедливо при любом типе роста, но не выяснив тип роста нельзя точно прогнозировать поведение Cw в зависимости от достигнутой массы. Выход из положения дает математическое описание роста. Средняя удельная скорость роста лишь в случае экспоненциального роста совпадает с удельной скоростью роста в момент времени t 1 или t 2. Для других типов роста средняя удельная скорость роста не передает изменение удельной скорости роста на интервале времени (t 1, t 2). Различия между средней удельной скорости роста за интервал времени (t 1, t 2) и удельными скоростями в моменты времени t 1 или t 2 возрастают с увеличением этого интервала.
Экспоненциальный рост. В тех случаях, когда рост животных не имеет ограничений, он происходит с постоянной удельной скоростью. Такой простейший тип роста животных называют экспоненциальным, т.к. изменение их размеров (массы) может быть передано экспоненциальной функцией времени:
Р= 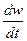 = Cw ∙ w
= Cw ∙ w
и
Cw = 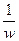 = const
= const
т.е. если известно, что рост экспоненциальный, то избавляемся от необходимости многократного расчета Cw, поскольку продукция (скорость роста) в этом случае постоянна в ходе роста.
Параболический рост. У многих животных удельная скорость энергетического обмена не остается постоянной в процессе их развития, а снижается по мере увеличения массы тела пропорционально массе в некоторой степени (W-n), обычно равной примерно 0,25 (n –константа уравнения зависимости интенсивности обмена у животных от их массы). Соответственно по мере роста животного и увеличения его массы снижается и удельная скорость роста
Р= = a∙ w b
где a и b – коэффициенты, получаем
Cw = a∙ w 1-b
Здесь qw снижается пропорционально w 1-b. Зная a и b, легко рассчитать Cw для любого w.
Поскольку животные резко различаются по массе и продолжительности периода роста, важное значение приобретает выбор подходящего сравнительного показателя, характеризующего скорость роста. Один из наилучших – удельная скорость роста Cw, вычисляемое по уравнению [1].
Уравнение параболического роста можно использовать для описания роста на отдельных этапах развития животных различного систематического положения. В ряде случаев оно хорошо описывает рост животных на протяжении всего их онтогенеза. Это относится к таким животным, рост которых по достижении определенного возраста прекращается. Но они продолжают жить еще некоторое время. Среди ракообразных такой тип роста свойственен веслоногим рачкам, у которых рост тела прекращается при пятой (последней) линьке копеподитов.
Асимптотический, или S-образный, рост. В онтогенезе подавляющего большинства водных животных различного систематического положения удельная скорость роста с увеличением массы тела уменьшается больше, чем удельная скорость энергетического обмена, пропорциональная W-n. Скорость роста таких животных в начальный период растет, достигая максимума, которому соответствует точка перегиба на кривой роста, а затем понижается.
Подобный характер роста может быть описан с помощью различных математических приемов, причем наиболее широко используется уравнение Берталанфи.
В наиболее общей форме уравнение Берталанфи выглядит следующим образом:
Р= = a1 w b1 – a2 w b2
где w – масса особи; - скорость роста массы (прирост); a1, a2, b1, b2 – коэффициенты. По Л. Берталанфи, в правой части уравнения первый член a1 w b1 отражает «анаболизм», величина которого пропорциональна поверхности тела животного, потому b1= 2/3. Второй член a2 w b2 отражает «катаболизм», пропорциональный массе тела (b2= 1). Следовательно, уравнение Берталанфи обычно используется в виде
|
|
|
|
|
Дата добавления: 2014-01-13; Просмотров: 735; Нарушение авторских прав?; Мы поможем в написании вашей работы!