КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Ферменты генетической инженерии
|
|
|
|
Рестриктазы
Рестриктазы (рестрицирующие эндонуклеазы, эндонуклеазы рестрикции) - это ферменты, узнающие и атакующие определенные последовательности нуклеотидов в молекуле ДНК (сайты рестрикции).
Еще в 1953 году было обнаружено, что ДНК определенного штамма E. Coli., введенная в клетки другого штамма (например, ДНК штамма В - в клетки штамма С) не проявляет, как правило, генетической активности, так как быстро расщепляется на мелкие фрагменты. В 1966 году было показано, что это явление связано со специфической модификацией хозяйской ДНК - она содержит несколько метилированных оснований, отсутствующих в немодифицированной ДНК, причем метилирование (добавление к основанию метильной группы) происходит уже после завершения репликации. Бактерия способна отличить свою собственную ДНК от любой вторгающейся «чужеродной» именно по типу ее модификации. За «метку» отвечают метилирующие ферменты модификации, так называемые ДНК-метилазы. Различие в модификации делает чужеродную ДНК чувствительной к действию рестрицирующих ферментов, которые узнают отсутствие метильных групп в соответствующих сайтах.
Системы рестрикции и модификации широко распространены у бактерий; их существование играет важную роль в защите резидентной ДНК от загрязнения последовательностями чужеродного происхождения. Рестриктаза, которая расщепляла неметилированную ДНК, была выделена в 1968 г. Мезельсоном и Юанем. Этот фермент был высокоспецифичен по отношению к определенной последовательности ДНК, но расщеплял молекулы неспецифически, в другом месте, на некотором удалении от участка узнавания. Вскоре, в 1970 г. Смит и Вилькокс выделили из Haemophilus influenzae первую рестриктазу, которая расщепляла строго определенную последовательность ДНК (Hind III). Поскольку разные бактерии по-разному метят свою ДНК, то и рестриктазы должны узнавать разные последовательности. И действительно, с тех пор выделены рестриктазы, узнающие более 150 сайтов рестрикции (мест расщепления ДНК).
Общепринято термины "рестриктаза", "эндонуклеаза рестрикции" и "сайт специфическая эндодезоксирибонуклеаза" считать синонимами.
Все рестрикционные эндонуклеазы бактерий узнают специфические, довольно короткие последовательности ДНК и связываются с ними. Этот процесс сопровождается разрезанием молекулы ДНК либо в самом сайте 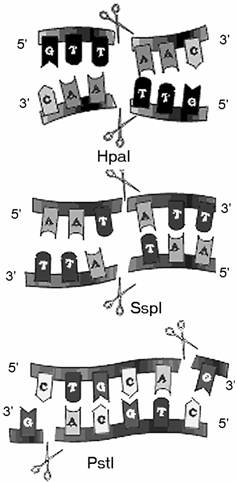 узнавания, либо в каком-то другом, что определяется типом фермента. Наряду с рестрикционной активностью бактериальный штамм обладает способностью метилировать ДНК; для этого процесса характерна такая же специфичность в отношении последовательностей ДНК, как и для рестрикции. Метилаза добавляет метильные группы к адениновым или цитозиновым остаткам в том же сайте, в котором связывается рестрикционный фермент. В результате метилирования сайт становится устойчивым к рестрикции. Следовательно, метилирование защищает ДНК от разрезания.
узнавания, либо в каком-то другом, что определяется типом фермента. Наряду с рестрикционной активностью бактериальный штамм обладает способностью метилировать ДНК; для этого процесса характерна такая же специфичность в отношении последовательностей ДНК, как и для рестрикции. Метилаза добавляет метильные группы к адениновым или цитозиновым остаткам в том же сайте, в котором связывается рестрикционный фермент. В результате метилирования сайт становится устойчивым к рестрикции. Следовательно, метилирование защищает ДНК от разрезания.
Классификация рестриктаз
Различают 3 основных класса рестриктаз: 1, 2 и 3.
Все рестриктазы узнают на двуспиральной ДНК строго определенные последовательности, но рестриктазы 1-го класса осуществляют разрывы в произвольных точках молекулы ДНК, а рестриктазы 2-го и 3-го классов узнают и расщепляют ДНК в строго определенных точках внутри сайтов узнавания или на фиксированном от них расстоянии.
Ферменты типов 1 и 3 имеют сложную субъединичную структуру и обладают двумя типами активностей - модифицирующей (метилирующей) и АТФ-зависимой эндонуклеазной.
Ферменты второго класса состоят из 2 отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы, поэтому в генной инженерии используются исключительно ферменты 2-го класса. Они нуждаются в ионах магния в качестве кофакторов.
В настоящее время выделено более 500 рестриктаз класса 2, однако среди ферментов, выделенных из различных микроорганизмов, встречаются такие, которые узнают на ДНК одни и те же последовательности. Такие пары или группы называют изошизомерами. Различают истинную изошизомерию, когда ферменты узнают одну и ту же последовательность нуклеотидов и разрывают ДНК в одних и тех же точках, и ложную, когда ферменты, узнавая один и тот же сайт на ДНК, производят разрывы в разных точках в пределах того же сайта. Большинство рестриктаз класса 2 узнают последовательности, содержащие от 4 до 6 нуклеотидных пар, поэтому рестриктазы делят на мелко- и крупнощепящие. Мелкощепящие рестриктазы узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем крупнощепящие, узнающие последовательность из шести нуклеотидных пар. Это связано с тем, что вероятность встречаемости определенной последовательности из четырех нуклеотидов гораздо выше, чем последовательности из шести нуклеотидов. К мелкощепящим относятся рестриктазы Hpa II и Alu (из Arthrobacter luteus), к крупнощепящим - Eco R I (из Escherichia coli) и Hind III. Если предположить, что участки узнавания рестриктаз распределены вдоль цепи ДНК случайно, то мишень для ферментов, узнающих последовательность (сайт) из четырех нуклеотидов, должна встречаться в среднем 1 раз через каждые 256 пар оснований, а для ферментов, узнающих шесть нуклеотидов, - через 4096 пар оснований. Если сайт рестрикции окажется внутри гена, то обработка ДНК-рестриктазой приведет к его инактивации. Вероятность такого события очень велика при обработке мелкощепящими рестриктазами и незначительна при применении крупнощепящих эндонуклеаз. Поэтому с целью получения неповрежденного гена расщепление проводят поочередно несколькими крупнощепящими рестриктазами, либо применяют прием "недорестрикции", т.е. рестрикцию проводят в таких условиях, когда происходит расщепление лишь в одном сайте.
Рестриктазы по-разному расщепляют ДНК (рис. 36). Одни вносят разрывы по оси симметрии узнаваемой последовательности (Hpa I, Ssp I). Другие - со сдвигом, со "ступенькой" (Pst I).
В первом случае образуются так называемые "тупые" концы, а во втором - "липкие", то есть фрагменты имеют на своих концах однонитевые взаимно комплементарные участки длиной в четыре нуклеотида. Такие фрагменты особенно удобны для создания рекомбинантных ДНК.
Обратная транскриптаза
Обратная транскриптаза используется для транскрипции м-РНК в комплементарную цепь ДНК. При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Наиболее детально изучена ревертаза ретровирусов птиц. Каждый вирион содержит около 50 молекул этого фермента. Обратная транскриптаза состоит из двух субъединиц — a (65 кДа) и b (95 кДа), присутствующих в эквимолярном количестве. Обратная транскриптаза обладает, по крайней мере, тремя ферментативными активностями:
1) ДНК-полимеразной, использующей в качестве матрицы как РНК, так и ДНК;
2) активностью РНКазы Н, гидролизующей РНК в составе гибрида РНК—ДНК, но не одно- или двухцепочечную РНК;
3) ДНК-эндонуклеазной активностью.
Первые две активности необходимы для синтеза вирусной ДНК, а эндонуклеаза, по-видимому, важна для интеграции вирусной ДНК в геном клетки-хозяина. Очищенная обратная транскриптаза синтезирует ДНК как на РНК-, так и на ДНК-матрицах. Чтобы начать синтез, ревертазе, как и другим полимеразам, необходим короткий двухцепочечный участок (праймер). Праймером может служить одноцепочечный сегмент как РНК, так и ДНК, которые в процессе реакции оказываются ковалентно связанными с новосинтезированной цепью ДНК.
Обратную транскриптазу преимущественно используют для транскрипции матричной РНК в комплементарную ДНК (кДНК).
Лигазы
В 1961 г. Мезельсон и Вейгл на примере фага l показали, что рекомбинация включает разрыв и последующее воссоединение молекул ДНК. Это положило начало поискам фермента, участвующего в сшивании фрагментов ДНК. В 1967 году такой фермент был найден и получил название ДНК-лигаза. Он катализирует синтез фосфодиэфирной связи в 2-х цепочечной молекуле нуклеиновой кислоты.
Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя связь между остатками сахаров. ДНК-лигазы абсолютно необходимы в процессах репарации ДНК, в процессах репликации - при удвоении цепи ДНК.
Существует 2 типа ДНК-лигаз, отличающихся по потребностям в кофакторах и способу действия. ДНК-лигаза E. coli в качестве кофактора использует дифосфопиридиннуклеотид, а лигаза фага Т4 - АТФ в присутствии Mg2+. Лигаза фага Т4 более универсальна, так как помимо лигирования липких концов способна катализировать реакцию воссоединения двухцепочечных фрагментов ДНК с тупыми концами. Она используется чаще.
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 2449; Нарушение авторских прав?; Мы поможем в написании вашей работы!