КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Наземные биомы (экосистемы)
|
|
|
|
Классификация природных экосистем биосферы на ландшафтной основе
ПРИРОДНЫЕ ЭКОСИСТЕМЫ ЗЕМЛИ КАК ХОРОЛОГИЧЕСКИЕ ЕДИНИЦЫ БИОСФЕРЫ
Классификации природных систем биосферы базируются на ландшафтном подходе, так как экосистемы – неотъемлемая часть природных географических ландшафтов, образующих географическую (ландшафтную) оболочку Земли. Биогеоценозы (экосистемы) образуют на поверхности Земли так называемую биогеосферу, являющуюся основой биосферы, которую В. И. Вернадский называл «пленкой жизни», а В. Н. Сукачев – «биогеоценотическим покровом».
«Биогеоценотический покров» В. Н. Сукачева – это не что иное, как ряд природных экосистем, представляющих собой пространственные (хорологические) единицы (части, элементы) биосферы. Эти единицы, как правило, совпадают своими границами с ландшафтными элементами географической оболочки Земли.
Ландшафт – природный географический комплекс, в котором все основные компоненты (верхние горизонты литосферы, рельеф, климат, воды, почвы, биота) находятся в сложном взаимодействии, образуя однородную по условиям развития единую систему.
Ландшафтный подход в экологии имеет, прежде всего, большое значение для целей природопользования. По происхождению выделяют два основных типа ландшафтов – природный и антропогенный.
Природный ландшафт формируется исключительно под влиянием природных факторов и не преобразован хозяйственной деятельностью человека. Изначально выделяли следующие природные ландшафты:
– геохимический – обозначает участок, выделенный на основе единства состава и количества химических элементов и соединений. Интенсивность их накопления в ландшафте или, напротив, скорость самоочищения ландшафта могут служить показателями его устойчивости по отношению к антропогенным воздействиям;
– элементарный ландшафт обозначает участок, сложенный определенными породами, находящимися на одном элементе, рельефа, в равных условиях залегания грунтовых вод, с одинаковым характером растительных ассоциаций и одним типом почв;
– охраняемый ландшафт, на котором в установленном порядке регламентированы или запрещены все или отдельные виды хозяйственной деятельности.
Однако, как считают многие ученые, сейчас на суше преобладают антропогенные ландшафты или, во всяком случае, по распространенности они равны природным.
Антропогенный ландшафт – это бывший природный ландшафт, преобразованный хозяйственной деятельностью настолько что изменена связь его природных компонентов. Сюда относятся ландшафты:
– агрокультурный (сельскохозяйственный) – растительность которого в значительной степени заменена посевами и посадками сельскохозяйственных и садовых культур;
– техногенный, структура которого обусловлена техногенной деятельностью человека, связанной с использованием мощных технических средств (нарушение земель, загрязнение промышленными выбросами и т. п.); сюда же входит ландшафт индустриальный, образующийся в результате воздействия на среду крупных промышленных комплексов;
– городской (урбанистический) – с постройками, улицами и парками.
Границы географической (ландшафтной) оболочки Земли совпадают с границами биосферы, но поскольку в географическую оболочку входят и участки, где нет жизни, можно условно принимать, что биосфера входит в ее состав. Фактически же – это неразрывное единство, о чем свидетельствует и ландшафтный подход при выделении типов природных экосистем. Одним из таких примеров служит классификация по Р. X. Уиттекеру, использованная им при оценке продуктивности экосистем земного шара (табл. 7.1).
Таблица 7.1 Первичная биологическая продуктивность экосистем земного шара (по Р. X. Уиттекеру, 1980)
| Типы экосистем | Площадь 106 км2 | Чистая первичная продукция, г/м2 | Общая чистая продукция, 109 т/год | |
| Колебания | В среднем | |||
| Влажные тропические леса | 1000–3500 | 37,4 | ||
| Тропические сезонные леса | 7,5 | 1000–2500 | 12,0 | |
| Вечнозеленые леса умеренного пояса | 5,0 | 600-2500 | 6,5 | |
| Листопадные леса умеренного пояса | 7,0 | 600-2500 | 8,4 | |
| Бореальные леса (тайга) | 12,0 | 400–2000 | 9,6 | |
| Лесо-кустарниковые сообщества | 8,5 | 250-1200 | 6,0 | |
| Саванны | 15,0 | 200-2000 | 13,5 | |
| Лугостепи умеренного пояса | 9,0 | 200–1500 | 5,4 | |
| Тундра и высокогорье | 8,0 | 10–400 | 1,1 | |
| Пустыни и полупустыни | 18,0 | 10–250 | 1,6 | |
| Экстрем, пустыни, скалы, пески и др. | 24,0 | 0-10 | 0,07 | |
| Культивируемые земли | 17,0 | 100–3500 | 9,1 | |
| Болота и марши | 2,0 | 800-3500 | 4,0 | |
| Озера и реки | 2,0 | 100-1500 | 0,5 | |
| Материковые экосистемы в целом: | 149,0 | 0-3500 | ||
| Открытый океан | 332,0 | 2-400 | 41,5 | |
| Зоны апвеллинга | 0,4 | 400-1000 | 0,2 | |
| Континентальный шельф | 26,6 | 200-600 | 9,6 | |
| Заросли водорослей и рифы | 0,6 | 500-4000 | 1,6 | |
| Речные дельты (эстуарии) | 1,4 | 200-3500 | 2,1 | |
| Морские экосистемы в целом: | 361,0 | 2–4000 | ||
| Средняя и общая продуктивность биосферы | 510,0 | 0–4000 |
Главный источник энергии для ландшафтной оболочки, как и для бисферы, – солнечная радиациия. Для биосферы солнечная энергия – это прежде всего «движитель» биогеохимических циклов биофильных элементов и главный компонент фотосинтеза – источника первичной продукции. Как видно из табл. 7.1, продуктивность биосферы складывается из продуктивности различных природных экосистем (одновременно и энергий ландшафтов).
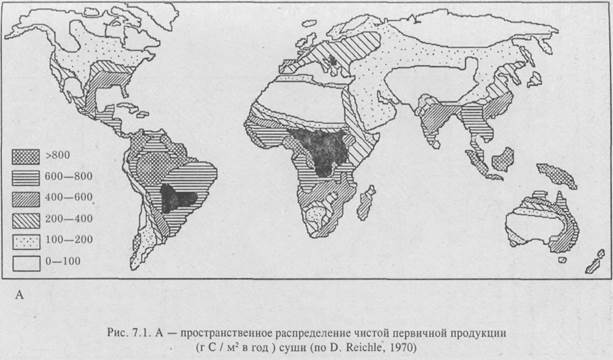
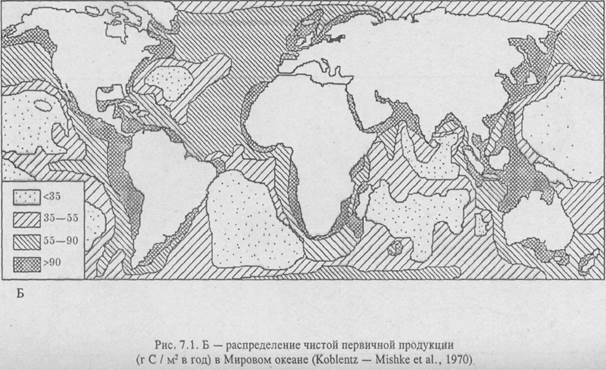
Но энергия Солнца, обеспечивая эту продуктивность, составляет лишь 2–3% от всей его энергии, достигшей поверхности Земли. Остальная солнечная энергия расходуется на абиотическую среду, если не считать достаточно активное участие ее в процессах физико-химического разложения, опада и др. Но абиотические факторы определяют вместе с биотическими эволюционное развитие организмов и гомеостаз экосистем. В свою очередь – растительный и животный мир – столь мощные природные компоненты, что могут влиять на окружающую среду и «переделать ее под себя», создавая определенную микросреду (микроклимат). Все это свидетельствует о том, что живая природа существует в едином энергетическом поле всего ландшафта. Об этом говорит и распределение первичной продукции, на суше и в океане (рис. 7.1; Бигон и др., 1989).
Как видно из рис. 7.1, продуктивность различных типов экосистем далеко не одинакова и занимают они разные по величине территории на планете. Различия в продуктивности связаны с климатической зональностью, характером среды обитания (суша, вода), с влиянием экологических факторов локального порядка их. п., сведения о которых излагаются ниже при характеристике природных экосистем как хорологических единиц биосферы, классифицированных на принципах так называемого биомного подхода. По Ю. Одуму (1986), биом – «крупная региональная и субконтинентальная экосистема, характеризующаяся каким-либо основным типом растительности или другой характерной особенностью ландшафта».
Опираясь на эти представления, Ю. Одум предложил следующую классификацию природных экосистем биосферы (на рис. 7.2 – мировое распределение биомов):
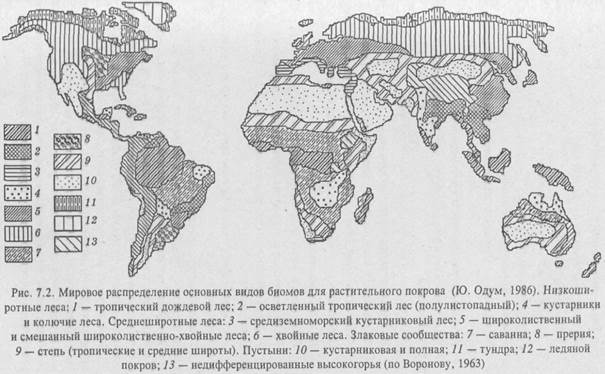
I. Наземные биомы.
Тундра: арктическая и альпийская.
Бореальные хвойные леса.
Листопадный лес умеренной зоны.
Степь умеренной зоны.
Тропические степи и саванны.
Чапарраль – районы с дождливой зимой и засушливым летом.
Пустыня: травянистая и кустарниковая.
Полувечнозеленый тропический лес: выраженный влажный и сухой сезоны.
Вечнозеленый тропический дождевой лес.
И. Типы пресноводных экосистем
Лентические (лат. lentes – спокойный): озера, пруды и т. д.
Лотические (лат. lotus – омывающий): реки, ручьи, родники.
Заболоченные угодья: болота и болотистые леса.
III. Типы морских экосистем
Открытый океан (пелагическая).
Воды континентального шельфа (прибрежные воды).
Районы апвеллинга (плодородные районы с продуктивным рыболовством).
Эстуарии (прибрежные бухты, проливы, устья рек, соленые марши и т. д.
Границы распространения биомов определяются ландшафтными компонентами материков, в названии, как правило, доминирующая растительность (лесной, кустарниковый и т. п.). В водных экосистемах растительные организмы не доминируют, поэтому за основу взяты физические признаки среды обитания («стоячая», «текучая» вода, открытый океан и т. п.).
Как явствует из вышесказанного, биом – это экосистема, которая совпадает своими границами с ландшафтами регионального уровня (рис. 7.2). Он состоит из тех же компонентов, что и ландшафт, но главный компонент его – биота, и основное внимание здесь уделяется процессам, создающим органическое вещество, и биохимическому круговороту веществ.
§
Стабильная экосистема характеризуется равновесным состоянием взаимосвязей между живыми организмами и окружающей физической средой. Всеобщий гомеостаз такой системы позволяет ей противостоять внешнему воздействию в довольно широком диапазоне толерантности по отношению, например, к климатическим факторам, тогда говорят об экосистеме климатического климакса.
Климатический климакс является результатом длительной сукцессионной серии экосистемы в данных климатических условиях. Но как известно, для Земли характерна климатическая зональность, а отсюда и зональность наземных экосистем, климаксная стадия которых будет определяться конкретными климатическими факторами соответствующей зоны.
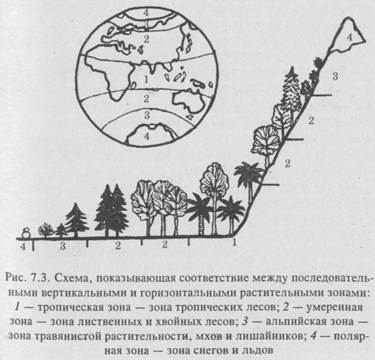
Известно, что кроме горизонтальной зональности (рис. 7.3) климата в масштабе всего земного шара, наблюдается еще и вертикальная, или высотная, зональность в горных системах. У подножия горных систем климат соответствует данной общегеографической зональности, при движении вверх, в горы пояса будут меняться, как при движении с юга на север. «Югом» будет являться климат подножия горной системы (рис. 7.3).
Тундры характеризуются суровыми условиями для произрастания: вегетационный период всего 2–2,5 месяца, осадков мало – 200–300 мм, сильные ветры и ночью температура падает ниже 0°С даже летом, плюс к этому – вечная мерзлота на глубине в несколько десятков сантиметров летом, а зимой – оттаивающий летом слой промерзает полностью. Но длительный фотопериод и низкая испаряемость значительно снижают лимитирующее действие влажности и света.
В тундре отсутствуют деревья и преобладают мхи и лишайники. Кустарники многолетние зимнезеленые для более полного использования светлой части года, подушкообразной и стелющейся форм (кассиопея, брусника, вероника и др.) и таких же форм карликовые растения с опадающими листьями (черника, карликовая береза и т. п.).
Продуктивность наземных экосистем тундры значительно ниже ряда других систем, но вместе с океаном они способны прокормить перелетных птиц, насекомых, северных оленей, овцебыков, медведей, волков, песцов и др.
Выше границы леса, в высоких горах есть небольшие тундровые зоны, которые являются экологически сходными районами с вышеописанной тундрой.
Бореальные хвойные леса распространены в северной части умеренной климатической зоны (рис. 7.2). Это хвойные леса северной части умеренного пояса северного полушария с суровыми зимними температурами – тайга. Таежные сообщества представлены темнохвойными породами деревьев – ель, пихта, сибирская кедровая сосна (сибирский кедр) и светлохвойными – лиственницей и сосной (преимущественно на песчаных и супесчаных почвах).
Строение сообществ темнохвойных лесов довольно простое: два-три древесных яруса, ярусы моховой, травяной или травяно-кустарниковый. Затенение значительно, лесная подстилка разлагается медленно.
Темнохвойные леса отличаются особой микросредой – нет ветра, температура выше, чем на открытом месте, много снега зимой, что помогает выживать животному населению в суровую зиму.
Самые крупные животные в тайге: из хищников – медведь, волк; из травоядных – лось. Большое значение в хвойных лесах имеет семенной фонд и хвоя: семенами питаются птицы, белки, бурундуки и другие мелкие грызуны, а хвоей – насекомые.
Хвойные леса – самые крупные в мире поставщики лесоматериалов. Они весьма продуктивны, несмотря на низкую температуру в течение полугода, так как сплошной зеленый покров, содержащий хлорофилл, сохраняется круглый год (см. табл. 7.1, рис. 7.1).
Листопадные леса умеренной зоны (широколиственные леса), расположенные южнее тайги, в отличие от нее, не имеют сплошного распространения (рис. 7.2). Произрастают они в условиях более мягкого климата, с осадками от 700 до 1500 мм/г, с умеренными температурами и четко выраженными сезонами. В основном в листопадных лесах среди древесной флоры доминируют бук и дуб. Благодаря опаду листьев формируется мощная лесная подстилка, позволяющая перезимовать многим беспозвоночным животным.
Ярусная структура широколиственных лесов намного сложнее, чем у хвойных: до трех ярусов деревьев (дубравы), двух ярусов кустарников и двух-трех ярусов трав. Раскидистая крона, высокая дуплистость деревьев и хорошо выраженная ярусность широколиственных лесов позволяют птицам занимать свои экологические ниши на различных уровнях.
Среди насекомых много вредителей, которые наносят большой вред лесам. Крупные животные в листопадных лесах, в принципе, те же, что и в тайге: лоси, медведи, рыси, лисицы и др., разнообразна и богата орнитофауна.
Листопадные леса – это те районы суши, где человеческая цивилизация получила наибольшее развитие. Поэтому трудно сейчас найти широколиственные нетронутые леса. Большая их часть заменена культурными сообществами.
Биомасса широколиственных лесов близка к биомассе южной тайги (см. табл. 7.1, рис. 7.1).
Степи умеренной зоны – открытые пространства между лесами и пустынями с количеством осадков от 250 до 750 мм/г. Они занимают обширные пространства в Евразии, в Северной Америке (прерии), на юге Южной Америки (пампасы), в Австралии, Новой Зеландии (туссоки) (рис. 7.2).
Растительность в степях преимущественно ксерофильного облика. Преобладают дерновинные злаки (дерновины, образуются вследствие скученности стеблей). Много в степи эфемеров, после отмирания наземных частей которых остаются клубни, луковицы, подземные корневища. И наконец, для степей характерны кустарники, которые поедаются животными.
Среди животных в степях распространен парный и – реже – колониальный образ жизни. Здесь много парных животных (суслики, сурки, полевки и др.). Животные, которые живут не в парах, образуют стада. Главную роль в биоценозе степей играют копытные – сайгаки, ранее дикая лошадь – тарпан. При умеренном выпасе животные копытами разбивают скопления мертвой листвы на поверхности почвы, что способствует дальнейшему росту трав. При перевыпасе происходит деградация степной растительности и возникает, в конечном итоге, так называемый «толок», когда практически исчезают все многолетние травы, а следом за «толоком» – опустынивание степей – растительность сменяется малосъедобными полынями и другими еще более ксерофильными формами.
Почвы степей достаточно резко отличаются от лесных почв и, прежде всего, высоким содержанием гумуса – в пять-десять раз выше. Злаки, по сравнению с деревьями, живут недолго и в почву попадает большое количество органики в виде гумуса, так как гумификация идет быстро в сухом климате, а минерализация очень медленно. Так возникают самые плодородные почвы – черноземы. На них вырастает наиболее высокая чистая первичная продукция, или урожай, культурных злаков – пшеницы, кукурузы и т. д.
В луговых степях России биомасса – 2500 ц/га, в сухих степях – 1000 ц/га. Продуктивность ксерофильных сообществ 100–200 ц/га, при возрастании аридности – 50–100 ц/га (Воронов, 1988).
Большая часть степей в настоящее время занята под посевы зерновых культур, культурными пастбищами или древесной растительностью, сохраненной или культивированной человеком.
Пустыня травянистая и кустарниковая наибольшие площади занимает в Азии, Африке, Австралии, Северной и Южной Америках, встречается она и в Европе (чаще полупустыня) (см. рис. 7.2). Главный критерий пустыни – выпадение осадков менее 200–250 мм/г, а испарение с открытой водной поверхности более 1000 мм/г.
Почвы пустынь – сероземы и светло-бурые. Пустыни обычно подразделяются по породам, на которых они сформировались: глинистые, солончаковые, песчаные, каменистые.
Растения в пустынях представлены весьма ксерофильными травами и полукустарниками, суккулентами, а также множеством эфемеров, которые используют только влажные периоды. Растительность разрежена, из-за чего травоядные животные существуют небольшими группами, парами и в одиночку. Стада образуют лишь животные, способные быстро находить новые участки с кормами (антилопы, некоторые птицы).
Животные пустыни по-разному адаптированы к нехватке воды: обладают особыми покровами, выделяют сухие экскременты и т. д. Они могут образовывать и сохранять метаболическую воду. Верблюды же приспособлены к повышенной температуре тела, к высокой степени дегидратации тканей, смертельной для других животных.
Земледелие в пустынях невозможно без орошения. При орошении, с ее изобилием солнечного света, пустыня может стать весьма продуктивной. По размерам фитомассы пустыни разнообразны (см. табл. 7.1, рис. 7.1).
Чапарраль – это территории с мягким, умеренным климатом. Количество осадков здесь 500–700 мм, но они выпадают в период теплой зимы. Обильные зимние дожди сменяются засушливым летом. Сообщества чапарраля состоят из деревьев (лавр, вечнозеленые дубы) и кустарников с желтыми толстыми вечнозелеными листьями. Они широко распространены в Средиземноморье, вдоль южного берега Австралии, в Калифорнии и Мексике (см. рис. 7.2).
В Австралии в лесах доминируют эвкалиптовые деревья и кустарники. Деревья широко интродуцируются в других местах этих биомов – в Калифорнии (США), в Колхидской низменности (Грузия).
Тропические степи и саванны – это обычно древесно-кустарниковый тип растительности теплых областей в центральной и Восточной Африке, в Южной Америке и Австралии, с осадками от 900 до 1500 мм/г. Температура здесь достаточно высока круглый год и сезонность определяется только распределением осадков – сезоны влажные (дождливые) и сухие (засушливые). Это создает своеобразные условия для существования фауны и флоры.
Деревья имеют часто толстую кору с мощным слоем пробки: баобабы, акации, пальмы, древовидные молочаи (экологические эквиваленты кактусов) и др. Травы представлены высокими труднопроходимыми для человека густыми злаками. В период засухи надземная часть злаков высыхает, листья деревьев опадают. Цветут деревья в конце сухого сезона, а с началом дождей распускается листва.
Саванна, в особенности африканская, не имеет себе равных по разнообразию и численности популяций копытных – антилопы, зебры, жирафы и др. На них охотятся такие хищники, как львы, гепарды и др.
Разнообразны птицы, среди которых есть крупные хищники и падальщики (грифы), а также самая крупная из птиц – африканский страус. Здесь множество рептилий – змей и ящериц, активных в засушливый период, а также насекомые, обилие которых приходится на дождливый сезон.
Среди насекомых много кровососущих – знаменитая муха цеце и др. В Южной Африке обитают насекомые, переносящие возбудителей тяжелых болезней, поражающих центральную нервную систему человека и животных, и других опасных «тропических» болезней.
Полувечнозеленые сезонные (листопадные) тропические леса распространены в областях с осадками 800–1300 мм/г с продолжительным сухим периодом в четыре – шесть месяцев в году. Леса характерны для тропической части Азии,
Центральной Америки (см. рис. 7.2). Характерно, что в них доминируют деревья верхнего яруса, которые сбрасывают листья в сухой сезон, соответствующий зимнему периоду. Нижний ярус представлен уже большей частью вечнозелеными деревьями и кустарниками. Из вечнозеленых деревьев в этих лесах хорошо известна пальма.
Вечнозеленые тропические дождевые леса расположены вдоль экватора, в зоне, где 2000–2500 мм/г осадков при достаточно равномерном распределении их по месяцам. Дождевые леса расположены в трех основных областях: 1) крупнейший сплошной массив в бассейне Амазонки и Ориноко в Южной Америке; 2) в бассейнах рек Конго, Нигера и Замбези в Африке и на острове Мадагаскар; 3) Индо-Малайской и островов Борнео – Новая Гвинея (рис. 7.2). Годовой ход температур в этих областях достаточно ровный и в ряде случаев снижает сезонные ритмы вообще или сглаживает их.
В дождевых тропических лесах деревья образуют три яруса: 1) редкие высокие деревья образуют верхний ярус над общим уровнем полога; 2) полог, образующий сплошной вечнозеленый покров на высоте 25–35 м; 3) нижний ярус, который четко проявляется как густой лес лишь в местах просвета в пологе. Травянистая растительность и кустарники практически отсутствуют. Но зато большое количество лиан и эпифитов. Видовое разнообразие растений очень велико – на нескольких гектарах можно встретить столько видов, сколько нет во флоре всей Европы (Ю. Одум, 1986). Число видов деревьев по разным учетам различно, но, видимо, достигает 170 и более, хотя трав – не более 20 видов. Количество видов межъярусных растений (лианы, эпифиты и др.) вместе с травами насчитывают 200–300 и более.
Влажные тропические леса – это достаточно древние климаксные экосистемы, в которых круговорот питательных веществ доведен до совершенства – они мало теряются и немедленно поступают в биологический круговорот, осуществляемый мутуалистическими организмами и неглубокими, большей частью воздушными, с мощной микоризой, корнями деревьев. Именно благодаря этому на скудных почвах так пышно растут леса.
Не менее разнообразен, чем растительность, и животный мир этих лесов. Большая часть животных, в том числе и млекопитающих, существуют в верхних ярусах растительности. Разнообразие видов животных можно проиллюстрировать такими цифрами: на 15 км2 дождевого леса в Панаме насчитывается 20 000 видов насекомых, а на такой же территории на западе Европы их всего несколько сотен.
Из крупных животных тропических лесов назовем лишь некоторые, наиболее известные: обезьяны, ягуары, муравьед, ленивец, пума, человекообразные обезьяны, буйвол, индийский слон, павлин, попугаи, кондор, королевский гриф и многие др.
Для тропического леса характерна высокая скорость эволюции и видообразования. Многие виды вошли в состав более северных сообществ. Поэтому очень важно сохранить эти леса как «ресурс генов».
Влажные тропические леса обладают большой биомассой и самой высокой продуктивностью из биоценозов суши (см. табл. 7.1).
Чтобы лес восстанавливался до состояния климакса, требуется длительный суккцессионный цикл. Для ускорения процесса предлагается, например, вырубать его узкими просеками, оставляя растения, которые ценности для промышленности не представляют, не нарушая при этом запас биогенов в корневых подушках, и тогда обсеменение с незатронутых участков поможет быстро восстановить лес до первоначального вида.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 4509; Нарушение авторских прав?; Мы поможем в написании вашей работы!