КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Динамика популяций
|
|
|
|
Биотический потенциал. Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного виду. Понятие биотического потенциала введено в экологию в 1928г. Р. Чепменом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например за год или за весь жизненный цикл.
При расчетах его чаще всего выражают коэффициентом r и вычисляют как максимально возможный прирост популяции ∆N за отрезок времени Δ t, отнесенный к одной особи, при начальной численности популяции N0:

|
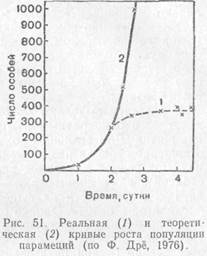
Величина биотического потенциала чрезвычайно различна у разных видов. Например, самка косули способна произвести за жизнь 10—15 козлят, трихина (Trichinella spiralis) — отложить 1,8 тыс. личинок, самка медоносной пчелы — 50 тыс. яиц, а луна-рыба — до 3 млрд. икринок. Биотический потенциал этих видов на самом деле еще больше, так как часть развивающихся яиц и зародышей погибает еще до рождения. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии.
Кривая, отражающая на графике подобный рост популяции, быстро увеличивает крутизну и уходит в бесконечность. Такая кривая носит название экспоненциальной. На логарифмической шкале подобная зависимость численности популяции от времени будет представлена прямой, а биотический потенциал г отразится ее наклоном по отношению к горизонтальной оси, который тем круче, чем больше величина г.
В природе биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяциях: r=b — d, где b — число родившихся, a d — число погибших особей в популяции за один и тот же период времени.
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (миграция).
Рождаемость. Это число новых особей, появляющихся в популяции за единицу времени в расчете на определенное число ее членов.
Различают абсолютную и удельную рождаемость. Первая характеризуется общим числом родившихся особей. Например, если в популяции северных оленей, насчитывающей 16 тыс. голов, за год появилось 2 тыс. оленят, то это число и выражает абсолютную рождаемость. Удельную вычисляют как среднее изменение численности на особь за определенный интервал времени, и в данном случае она составит 0,125, т. е. один новорожденный на 8 членов популяции за год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. Первые дают одну, вторые — несколько генераций за год. Например, у тлей за сезон насчитывается до 15 партеногенетических поколений. По числу периодов размножения в течение жизни различают моноциклические и полициклические виды. Моноцикличность свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые). Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни. Для величины рождаемости имеет значение также соотношение периода размножения и общей продолжительности жизни. Этот период для самок дрозофил составляет около 65%, для стадной саранчи шистоцерки—15%, а для поденок — всего от 0,5 до 1% длительности их существования. Большую роль играет плодовитость особей. Однако размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами. Среди рыб наибольшее количество икринок выметывают виды с пелагической икрой — сельди, тресковые, камбаловые и др. Например, сахалинская сельдь продуцирует 38—46 тыс. мелких, в доли миллиметра, икринок. У лососевых, зарывающих икру в грунт, развивается меньшее число яиц, но более крупных размеров. Средняя плодовитость амурской горбуши 1300—1500 икринок диаметром 4—6 мм. Наиболее крупная икра у акул и химер, до 6—8 см. Яйца этих рыб имеют к тому же плотную защитную оболочку. Количество их очень невелико — несколько штук на одну самку.
У видов, оберегающих и выкармливающих свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, чистиков и др.) до 20—25 (максимальное количество яиц у куриных, например, у серой куропатки).
Д. Лэк установил на примере птиц, что у них отбор благоприятствует не максимальной плодовитости, а наиболее эффективной, т. е. тому количеству яиц, при котором потомство оказывается наиболее жизнеспособным. На выкармливание птенцов птицы тратят огромное количество энергии. Мелкие птицы приносят пищу в гнездо сотни раз в сутки: горихвостка — более 200, большая синица — около 400, а крапивник — до 600 раз. Если величина кладки больше обычного предела, птенцы оказываются недокормленными и их жизнеспособность снижена.
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на более высокую плодовитость — одно из приспособлений, компенсирующих большую норму гибели в популяциях. Поэтому при высокой плодовитости общая размножаемость популяции может быть весьма низкой. В различных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих — зайцев, мышей, полевок —число детенышей в помете на границах ареала больше, чем в его центре.
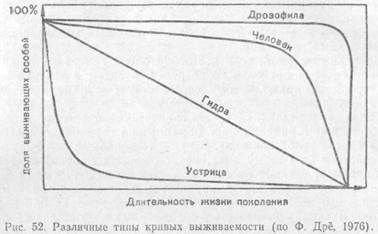
Смертность. Смертность в популяциях также зависит от многих причин: генетической и физиологической полноценности особей, влияния неблагоприятных физических условий среды, воздействия хищников, паразитов, болезней и т. п. На разных стадиях жизненного цикла каждого поколения эти факторы действуют с разной силой. На практике, когда необходимо проанализировать ход смертности в популяциях, составляют таблицы выживания (см. далее), где для каждой возрастной группы указываются эмпирически полученные данные, характеризующие гибель особей данного возраста. На основе таких таблиц составляют кривые выживания, позволяющие прогнозировать в сходных условиях состояние очередных генераций.
Таблица 1
Таблица выживания куропатки (по Швердпфегеру, 1968)
| Возраст, годы | Кол-во особей | Смертность, % | Доля самок в популяции | Средняя ожидаемая продолжит. жизни, годы | |
| живых к началу возраста | погибших в данном возрасте | ||||
| 0,50 | 1,0 | ||||
| 0,47 | 2,9 | ||||
| 0,46 | 2,7 | ||||
| 0,41 | 2,6 | ||||
| 0,32 | 2,5 | ||||
| 0,26 | 2,4 | ||||
| 0,23 | 2,3 | ||||
| 0,29 | 2,3 | ||||
| 0,27 | 2,3 | ||||
| 0,28 | 2,3 | ||||
| 0,20 | 2,0 | ||||
| 2,3 | |||||
| 1,5 | |||||
| - |
Идеальный случай, если все особи одной генерации доживают до биологически предельного возраста, а затем в течение короткого срока отмирают. Этому соответствует минимальная смертность популяции.
Кривая, описывающая такую зависимость численности поколений от времени, сначала идет параллельно горизонтальной оси, а затем круто загибается вниз. В природе подобный ход смертности не свойствен ни одному виду, но встречается приближение к нему, например, у таких насекомых с хорошо защищенными личинками, как жуки-златки, у крупных млекопитающих с высокой степенью выживаемости молодняка. К этому же типу близка кривая, описывающая смертность в современных популяциях человека. Средняя продолжительность жизни особи в таких популяциях высока и приближается к максимальной.
Наиболее часто встречающийся в природе вариант — повышенная гибель особей в ранний период жизни. Взрослые формы более защищены или выносливы. Кривая смертности в таких популяциях резко падает к горизонтальной оси в самом начале. Ее наклон отражает скорость убыли поколения. Так, у большинства рыб значительная часть популяции гибнет еще на стадии икры, очень велик также отсев мальков и лишь среди взрослых особей смертность становится ниже. У мышей и полевок, выкармливаемых матерью, гибель учащается после выхода молодых из гнезда, когда они приступают к самостоятельной жизни. Средняя продолжительность жизни особи намного меньше максимальной. У людей также на протяжении почти всей истории была высокой детская смертность, которая резко снизилась с успешным развитием медицины. Это привело к изменению типа кривой выживаемости в поколениях и стремительному увеличению численности населения Земли, получившему название «демографического взрыва».
Сравнительно редко наблюдается третий вариант изменения выживаемости поколений — относительно равномерный отсев из-за случайных причин на протяжении всего жизненного цикла, без резко выраженных критических периодов повышенной смертности. На графике это выражается прямой линией между начальной и нулевой численностью генерации. Такой ход смертности отмечен, например, у пресноводных гидр, содержащихся в аквариуме.
Расселение. Выселение особей из популяции или пополнение ее пришельцами — закономерное явление, основанное на одной из важнейших биологических черт вида — его расселительной способности. В каждой популяции часть особей регулярно покидает ее, пополняя соседние или заселяя новые, еще не занятые видом территории. Этот процесс называют часто дисперсией популяции. Расселение приводит к занятию новых биотопов, расширению общего ареала вида, его успеху в борьбе за существование.
Расселительные функции выполняются в определенный период жизненного цикла: у насекомых в основном на стадии имаго, у тироглифоидных клещей — специальными дейтонимфами-гипопусами, у большинства птиц и млекопитающих — подрастающим молодняком. У растений рассеиваются или разносятся семена и споры, сидячие животные распространяются посредством плавающих личинок или специальных поколений при метагенезе.
Каждый вид характеризуется своим темпом дисперсии. По подсчетам, у зайцев-беляков регулярно покидает места рождения примерно 1 % молодняка, тогда как в популяциях большой синицы в среднем лишь треть молодых остается в той местности, где она вывелись из яиц.
Дисперсия обычно не направлена, расселение особей происходит в самых разных направлениях от мест отрождения.
Расселительная дисперсия служит средством связи между популяциями. Она повышается при увеличении плотности населения.
В период депрессии численности, наоборот, усиливается поток вселенцев в популяцию. У оседлых животных с хорошо выраженными территориальными инстинктами агрессивное поведение по отношению к пришельцам в период низкой численности популяции ослабевает, и вселенцы занимают свободные участки.
Ряд популяций, занимающих малопригодные места обитания, часто не в состоянии поддерживать свою численность за счет размножения и могут сохраняться преимущественно за счет иммиграции. Такие популяции В. Н. Беклемишев называл зависимыми.
Расселительные перемещения, по Н. П. Наумову, приводят к обмену особей между популяциями, увеличивают единство и общую устойчивость вида, так как те адаптации, которые возникли в местных условиях, но имеют общее значение, могут постепенно распространяться в пределах всего видового ареала. Проникновение расселяющихся особей на не занятые еще видом территории, заселение их и образование новых популяций называют инвазией.
Темпы роста популяции. Графики роста числа членов любой природной популяции в новых для нее условиях сильно отличаются от экспоненты. Кривая после подъема разной степени крутизны поворачивает параллельно горизонтальной оси, чем знаменует установление некоторой предельной численности популяции, которая затем поддерживается в течение более или менее длительного времени. Такой ход кривой показывает, что в природе какие-то причины сдерживают чрезмерный рост популяции, не давая ей реализовать свой биотический потенциал, и ограничивают ее численность определенными пределами.
Изменения темпов роста популяции могут быть различными.
В одном случае темп роста с самого начала высок и постоянен независимо от нарастающей плотности, что соответствует лавинообразному, по экспоненте, увеличению численности популяции. При достижении же определенной плотности населения темп роста падает сразу до нуля.

Это означает, что популяция резко прекращает воспроизводство. В природе такой тип захвата среды встречается у видов, для которых жизненно важна скорость овладения ресурсами, например у нематод, клещей, некоторых насекомых, обитающих в быстро меняющих качество гниющих растительных остатках, кучках навоза копытных и т. п. Мелкие нематоды, населяющие конский и коровий навоз, могут осуществить свой жизненный цикл за несколько часов, тогда как близкие им виды, но обитающие в других, более постоянных условиях, развиваются от двух до трех недель. Скорость овладения средой имеет значение и для предупреждения конкуренции со стороны других претендентов. После массового размножения и быстрого потребления доступных ресурсов в популяциях подобных видов размножение прекращается и особи расселяются путем перелета на насекомых.
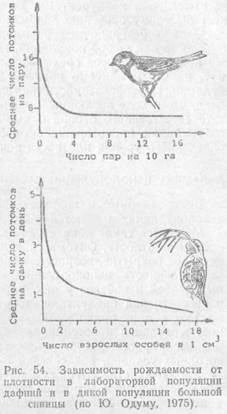
В другом случае изменения темпов популяций находятся в обратной зависимости прироста от плотности. Разреженная популяция в благоприятных условиях быстро наращивает свою численность, но чем больше становится в ней особей, тем меньше доля очередного приплода, пока скорость воспроизводства не сравняется со смертностью. Тогда темп роста популяции снижается до нуля, а общая численность стабилизируется в соответствии с доступными для популяции ресурсами. Такой характер становления новых популяций свойствен в основном видам, у которых успех в размножении не лимитируется обязательностью группового образа жизни, необходимостью встречи полов и другими причинами (например, у мелких партеногенетических ракообразных, растений с апомиксисом и т. д.). Однако у большинства видов наиболее высокий темп роста популяций отмечается лишь при определенной оптимальной плотности. Если популяция сильно разрежена, это затрудняет встречу полов, защиту молодняка, проявление зффекта группы у животных, опыление у растений, поэтому население растет в начале очень медленно.
|
Как известно, появление потомства прежде всего зависит от числа производителей — особей, находящихся в генеративном возрастном состоянии. Вначале рост числа производителей сопровождается медленным ростом, численности популяции, затем, в некотором интервале плотностей, зависимость выражена чрезвычайно резко, и даже небольшое увеличение доли производителей вызывает стремительный рост популяции, пока она не достигнет определенного уровня, который в последующем уже не меняется, сколько бы ни увеличивалась репродуктивная часть населения.
Такая зависимость была впервые предсказана французским математиком Верхюльстом в середине прошлого века для населения людей, а позднее доказана англичанином Перлом (1925) для популяций животных в среде, где пищевые ресурсы имеют определенный предел восполнимости.
Установление определенного уровня плотности популяций после некоторого периода роста вовсе не означает, что в популяциях больше не происходит количественных изменений. Напротив, любое население всегда динамично и постоянно подвержено колебаниям численности, однако размах всех суточных, сезонных и годовых изменений популяций все же намного меньше теоретически возможного, соответствующего реализации всего биотического потенциала. Колебания численности совершаются с разным размахом вокруг некоторой средней величины, которая и соответствует горизонтальной части кривой на графике роста и стабилизации численности популяции.
Высокий потенциал размножения играет большую роль в выживании видов. Популяции, сведенные к низкому уровню численности, могут быстро восстановиться при благоприятной перемене условий. Некоторые виды только массовым размножением, могут противостоять выеданию их различными потребителями или угрозе вытеснения конкурентами. Высокая размножаемость способствует быстрому освоению видом новых пространств.
Однако безграничное размножение таит в себе и большую опасность для любой популяции, так как может привести к быстрому подрыву ресурсов среды, нехватке пищи, убежищ, пространства и т. п., что неминуемо повлечет за собой общее ослабление популяции. Перенаселенность настолько неблагоприятна для любого вида, что в ходе эволюции у разных форм выработались в результате естественного отбора самые разнообразные механизмы, способствующие предотвращению избытка особей и поддержанию определенного уровня плотности популяций.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 897; Нарушение авторских прав?; Мы поможем в написании вашей работы!