КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Репликация ДНК. Для того чтобы начать серьезный разговор о репарации, необходимо вспомнить основные детали репликации ДНК
|
|
|
|
Для того чтобы начать серьезный разговор о репарации, необходимо вспомнить основные детали репликации ДНК. Важно понимать, что все процессы ДНК-метаболизма тесно связаны между собой и белки, вовлеченные в репликацию и рекомбинацию ДНК, способны принимать участие в различных процессах репарации. Для наглядности рассмотрим схему процесса репликации у прокариот, представленную на рис. 1.
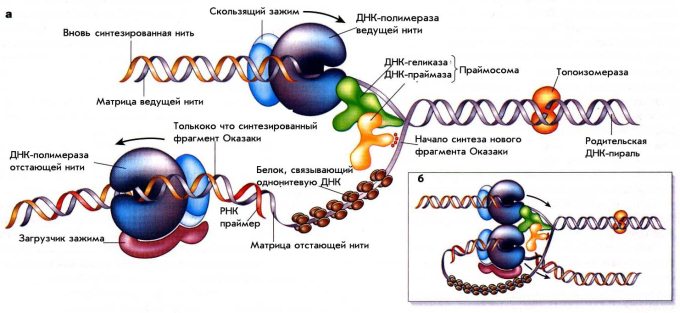
Рисунок 1. Схема репликации ДНК у E.coli. На панели а схематически показаны белки в Y– раскрученной вилке репликации ДНК, но в реальности вилка репликации свернута в трех измерениях и образует структуру, подобную той, что изображена на вставке б.
Сфокусируемся на схематическом изображении а. В репликативной вилке одновременно активны две молекулы ДНК-полимеразы. Одна движется постоянно, производя новую дочернюю молекулу ДНК на ведущей нити, тогда как другая производит длинную серию коротких фрагментов Оказаки на отстающей нити. Обе полимеразы «заякорены» на их матрице с помощью ассоциированных с ними белков в виде «скользящего зажима» (sliding clump) и «загрузчика зажима» (clump loader).
ДНК-геликаза, активированная энергией АТФ-гидролиза, быстро движется вдоль одной из матричных нитей (на рис. 1 – ведущая нить), раскрывая ДНК-спираль прямо перед репликативной вилкой. ДНК-геликаза предоставляет основания ДНК-спирали ДНК-полимеразе ведущей нити, чтобы та могла их копировать. Ферменты ДНК-топоизомеразы облегчают раскручивание спирали ДНК.
В дополнение к матрице, ДНК-полимераза нуждается в праймере, «затравке», существующей до начала собственно самого основного процесса синтеза ДНК, небольшой цепи ДНК или РНК с концом, к которому и присоединяется следующий нуклеотид. По этой причине ДНК-полимераза отстающей нити нуждается в действии фермента ДНК-праймазы перед тем, как она начнет синтез каждого нового фрагмента Оказаки. Праймаза производит очень короткую молекулу РНК (как РНК-праймер) в качестве 5’-конца каждого фрагмента Оказаки, к которому ДНК-полимераза и будет достраивать нуклеотиды. Наконец, однонитевые участки ДНК внутри вилки покрыты множеством копий SSB белка (связывающего однонитевую ДНК, single strand binding), занимая открытую область ДНК на нитях-матрицах с их подготовленными для копирования основаниями.
В представленной на вставке б схеме репликационной вилки показано, что ДНК-полимераза отстающей нити остается связанной с ДНК-полимеразой ведущей нити. Это позволяет ДНК-полимеразе отстающей нити оставаться внутри вилки после того, как завершен синтез очередного фрагмента Оказаки. В результате, одна и та же полимераза снова и снова участвует в синтезе большого числа фрагментов Оказаки, необходимых для образования новой цепи ДНК на отстающей нити.
В дополнение к перечисленной группе коровых (основных) белков, для репликации ДНК необходимы и другие белки, не представленные на рис. 1. Среди них можно описать группу белков – инициаторов репликации, необходимых для организации каждой новой репликативной вилки на ориджине репликации, белок РНКазу, удаляющий РНК-затравки фрагментов Оказаки, и ДНК-лигазу, сшивающая близлежащие фрагменты Оказаки друг с другом при формировании непрерывной нити ДНК.
RecQ-подобные геликазы вместе с топоизомеразой III также играют важную роль в репликации, а не только в других клеточных процессах, таких как гомологическая рекомбинация и контроль клеточного цикла.
Таким образом, мы видим, что процесс репликации очень сложен и может приводить к ошибкам в генетическом тексте, например, за счет подстановки неправильно спаренного или измененного основания. Одновременно процесс репликации является постоянным источником разрывов ДНК в клетке. Во-первых, всегда есть еще не соединившиеся фрагменты Оказаки. Во-вторых, при остановке движения репликативной вилки, чем бы она ни была вызвана, начинается процесс деградации ДНК и появляются ее свободные концы. В-третьих, топоизомераза I (ТорI) катализирует две основные реакции – разрезание и воссоединение однонитевой, нормально спаренной ДНК для релаксации ее суперскрученности при репликации или транскрипции. Множество эндогенных факторов действуют на эти две реакции разобщающее и приводят к образованию и накоплению ТорI-разрешающего комплекса, который является переходным к образованию двунитевых разрывов ДНК со всеми вытекающими последствиями. Таким образом, в течение процесса репликации структура ДНК может повреждаться и изменяться, а это уже само по себе является материалом для репарации.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 892; Нарушение авторских прав?; Мы поможем в написании вашей работы!