КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Система органов пищеварения. 10 страница
|
|
|
|
Как правило, в движении участвуют несколько мышц, которые по отношению друг к другу могут выступать как агонисты (действуют в одном направлении) либо антагонисты (действуют разнонаправленно). Рефлекторный акт возможен только при сопряженном, так называемом реципрокном торможении двигательных центров мышц-антагонистов. При ходьбе сгибание ноги сопровождается расслаблением разгибателей и, наоборот, при разгиба-нии тормозятся мышцы-сгибатели. Если бы этого не происходило, то возникла бы механи-ческая борьба мышц, судороги, а не приспособительные двигательные акты. При раздраже-нии чувствительного нерва, вызывающего сгибательный рефлекс, импульсы направляются к центрам мышц-сгибателей и через специальные вставочные нейроны (тормозные клетки Реншоу) — к центрам мышц-разгибателей. В первых вызывают процесс возбуждения, а во вторых — торможения. В ответ возникает координированный, согласованный рефлекторный акт — сгибательный рефлекс.
Взаимодействие процессов возбуждения и торможения — универсальный принцип, лежащий в основе деятельности нервной системы. Конечно, он реализуется не только на уровне сегментов спинного мозга. Вышестоящие отделы нервной системы осуществляют свое регуляторное влияние, вызывая процессы возбуждения и торможения нейронов нижестоящих отделов. Важно отметить: чем выше уровень животного, тем сильнее власть самых высших отделов центральной нервной системы, «тем в большей степени высший отдел является распорядителем и распределителем деятельности организма» (И.П.Павлов). У человека таким «распорядителем и распределителем» является кора больших полушарий головного мозга.
Каждый спинальный рефлекс имеет свое рецептивное поле и свою локализацию (место нахождения), свой уровень. Так, например, центр коленного рефлекса находится во II-IV поясничном сегменте; ахиллова — в V поясничном и I-II крестцовых сегментах; подош-венного — в I-II крестцовом, центр брюшных мышц — в VIII-XII грудных сегментах. Важнейшим жизненно важным центром спинного мозга является двигательный центр диафрагмы, расположенный в III-IV шейных сегментах. Повреждение его ведет к смерти вследствие остановки дыхания.
Кроме двигательных рефлекторных дуг, на уровне спинного мозга замыкаются вегета-тивные рефлекторные дуги, осуществляющие контроль за деятельностью внутренних орга-нов.
Межсегментарные рефлекторные связи. В спинном мозге помимо описанных выше рефлекторных дуг, ограниченных пределами одного или нескольких сегментов, действуют восходящие и нисходящие межсегментарные рефлекторные пути. Вставочными нейронами в них служат так называемые проприоспинальные нейроны, тела которых находятся в сером веществе спинного мозга, а аксоны поднимаются или спускаются на различные расстояния в составе проприоспинальных трактов белого вещества, никогда не покидая спинной мозг. Опыты с дегенерацией нервных структур (в которых полностью изолируются отдельные части спинного мозга) показали, что к проприоспинальным нейронам относится большинство его нервных клеток. Некоторые из них образуют независимые функциональные группы, ответственные за выполнение автоматических движений (автоматических программ спинного мозга). Межсегментарные рефлексы и эти программы способствуют координации движений, запускаемых на разных уровнях спинного мозга, в частности, передних и задних конечностей, конечностей и шеи.
Благодаря этим рефлексам и автоматическим программам спинной мозг способен обеспе-чивать сложные согласованные движения в ответ на соответствующий сигнал с периферии или от вышележащих отделов центральной нервной системы. Здесь можно говорить об интегративной (объединяющей) функции спинного мозга, хотя следует иметь в виду, что у высших позвоночных (в частности, у млекопитающих) возрастает регуляция спинальных функций высшими отделами центральной нервной системы (процесс энцефализации).
Спинальная локомоция. Обнаружено, что основные характеристики локомоции, т.е. пере-мещения человека или животного в окружающей среде при помощи координированных движений конечностей, запрограммированы на уровне спинного мозга. Болевое раздражение какой-либо конечности спинального животного вызывает рефлекторные движения всех четырех; если же такая стимуляция продолжается достаточно долго, могут возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся раздражению конечностей. Если такое животное поставить на тредмилл (бегущую дорожку), то при некоторых условиях оно будет совершать координированные шагательные движения, весьма сходные с естественными.
У спинального животного, анестезированного и парализованного кураре, в определенных условиях можно зарегистрировать ритмично чередующиеся залпы импульсов мотонейронов разгибателей и сгибателей, примерно соответствующие наблюдаемым при естественной ходьбе. Поскольку такая импульсация не сопровождается движениями, ее называют ложной локомоцией. Она обеспечивается пока еще не идентифицированными локомоторными центра-ми спинного мозга. По-видимому, для каждой конечности существует один такой центр. Активность центров координируется проприоспинальными системами и трактами, пере-секающими спинной мозг в пределах отдельных сегментов.
Предполагают, что у человека тоже есть спинальные локомоторные центры. По-видимому, их активация при раздражении кожи проявляется в виде шагательного рефлекса новорожденного. Однако по мере созревания центральной нервной системы вышестоящие отделы, очевидно, настолько подчиняют себе такие центры. что у взрослого человека они утрачивают способность к самостоятельной активности. Тем не менее, активизация локомоторных центров путем интенсивной тренировки лежит в основе различных методик восстановления ходьбы у больных с повреждением спинного мозга.
Таким образом, даже на уровне спинного мозга обеспечиваются запрограммированные (автоматические) двигательные акты. Подобные независимые от внешней стимуляции двигательные программы шире представлены в высших двигательных центрах. Некоторые из них (например, дыхание) врожденные, другие же (например, езда на велосипеде) приобретаются в процессе научения
Центральная нервная система. Белое вещество ЦНС состоит из длинных, покрытых белым миэлином, аксонов клеток и нейроглии. Тела этих клеток лежат в сером веществе или в ганглиях вне ЦНС. Будучи проводящей системой мозга, белое вещество осуществляет двусторонние связи между различными участками мозга, создавая таким образом ЦНС. В проводящих путях мозга волокна нервных клеток объединяются в пучки.
Таблица 11. Центральная нервная система
| Нервная система | Головной мозг | Спинной мозг | ||||||
| большие полушария | мозжечок | ствол | ||||||
| Состав и строение | Доли: лобная, те-менная, затылочная, две височные. Кора образована серым ве-ществом - телами нервных клеток. Тол-щина коры 1,5-3 мм Площадь коры - 2-2.5 тыс см2, она состоит из 14 млрд. тел нейронов. Белое ве-щество образовано нервными отростками | Серое вещество образует кору и ядра внутри мозжечка Состоит на двух полушарий, соеди-ненных мостом | Образован: 1. Промежуточным моз-гом 2. Средним мозгом 3. Мостом 4. Продолговатым моз-гом. Состоит из белого ве-щества, в толще нахо-дятся ядра серого ве-щества. Ствол переходит в спинной мозг | Цилиндрический тяж 42- 45 см длиной и около 1 см диаметром. Проходит в позвоночном канале. Внутри него находится спинно-мозговой канал, заполненный жидкостью. Серое вещество распо-ложено внутри, белое - снаружи. Переходит в ствол головного мозга, образуя единую систему | ||||
 Функции Функции
| Осуществляет выс-шую нервную дея-тельность (мышление, речь, вторая сигналь-ная система, память, воображение, способ-ность писать, читать) Связь с внешней сре-дой происходит с по-мощью анализаторов, находящихся в заты-лочной доле (зритель-ная зона), в височной доле (слуховая зона), вдоль центральной бо-розды (кожно-мышеч-ная зона) и на внут- | Регулирует и координирует дви-жения тела, мышеч-ный тонус Осущест-вляет безусловно-ре-флекторную деятель-ность (центры врож-денных рефлексов) | Связывает головной мозг со спинным в еди-ную центральную нерв-ную систему. В продолговатом мозге находятся центры: ды-хательный, пищевари-тельный, сердечно-со-судистый. Мост свя-зывает обе половины мозжечка. Средний мозг контролирует реакции на внешние раздражители, тонус (напряжение) мышц. | Функционирует под контролем головного мозга. Через него прохо-дят дуги безусловных (врожденных) рефлексов, осуществляющих возбуж-дение и торможение при движении. Проводящие пути белое вещество, соединяющее головной мозг со спинным; является проводником нервных импульсов. Регулирует работу внутренних орга-нов через периферическую нервную систему. Через | ||||
Продолжение таблицы 11
| Нервная система | Головной мозг | Спинной мозг | ||
| большие полушария | мозжечок | ствол | ||
| Функции
| ренней поверхности коры (вкусовая и обо-нятельная зоны). Регу-лирует работу всего организма через пери-ферическую нервную систему | Промежуточный мозг регулирует обмен веществ, температуру температуру тела, связывает рецепторы тела с корой больших полушарий | спинно-мозговые нервы осуществляется управ-ление произвольными движениями тела |
Головной мозг. Головной мозг состоит из больших полушарий (рис. 300-303) и ствола. Б ольшие полушария в глубине соединены большой спайкой — мозолистым телом. В них различают лобную, теменную, височную, затылочную доли и островок. В полушариях находятся боковые желудочки мозга, подкорковые ядра, внутренняя капсула. Доли мозга отделены друг от друга глубокими бороздами, среди которых наиболее выражены три глубоких борозды: центральная (роландова). отделяющая лобную долю от теменной, латеральная (сильвиева), ограничивающая лобную и теменную доли от височной и теменно-затылочная, проходящая по внутренней поверхности полушария и отделяющая теменную долю от затылочной. Наличие борозд и извилин значительно увеличивает общую площадь коры больших полушарий (до 2500 см), причем 2/3 поверхности находится в глубине борозд, а 1/3 — на поверхности полушарий. Головной мозг представляет собой расширенный передний конец спинного мозга. У человека это расширение настолько велико, что сходство со спинным мозгом в значительной степени замаскировано, но у низших животных структурное родство головного мозга со спинным ясно заметно.
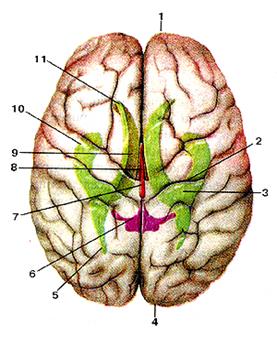
Рисунок 300. Большой мозг (cerebrum). Проекция боковых желудочков на поверхность полу-шарий большого мозга. Вид сверху. I - лобная доля; 2 - центральная борозда; З - боковой желудочек; 4 - затылочная доля; 5 - задний рог бокового желудочка; 6 - IV желудочек; 7 -водопровод мозга; 8 - III желудочек; 9 - центральная часть бокового желудочка; 10 - нижний рог бокового желудочка; 11 - передний рог бокового желудочка.
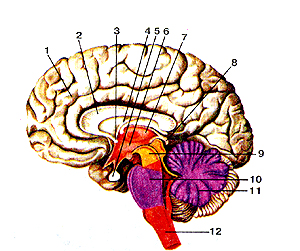
Рисунок 301. Головной мозг (cerebrum). Сагиттальный разрез. Вид с медиальной стороны.
I - полушарие большого мозга; 2 - мозолистое тело; 3 - передняя (белая) спайка; 4 - свод мозга; 5 - гипофиз; 6 - полость промежуточного мозга (III желудочек); 7 - таламус; 8 - эпифиз мозга; 9 - средний мозг; 10 - мост; 11 - мозжечок; 12 - продолговатый мозг.
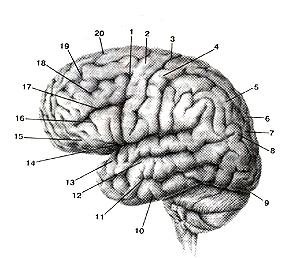
Рисунок 302. Верхне-латеральная поверхность полушария большого мозга. I - пред-центральная борозда; 2 - предцентральная извилина; 3 - центральная борозда; 4 - постц-ентральная извилина; 5 - верхняя теменная долька; 6 - внутритеменная борозда; 7 - нижняя теменная долька; 8 - угловая извилина; 9 - затылочной полюс; 10 - нижняя височная извилина; 11 - нижняя височная борозда; 12 - средняя височная извилина; 13 - верхняя височная извилина; 14 - латеральная борозда; 15 - глазничная часть; 16 - нижняя лобная извилина; 17 - нижняя лобная борозда; 18 - средняя лобная извилина; 19 - верхняя лобная борозда; 20 - верхняя лобная извилина.
Большие полушария мозга — самый передний (рис. 303) и наиболее крупный из отделов головного мозга — обладают совершенно иной функцией, заключающейся в регуляции приобретенных форм. поведения. В основе сложных психологических явлений сознания, умственной деятельности, памяти, понимания и истолкования ощущений лежит активность нейронов большого мозга. Значение больших полушарий у различных животных можно изучать, удаляя их хирургическим путем. Лягушка после такой операции ведет себя почти в точности так же, как нормальная лягушка, а голубь с удаленной корой больших полушарий способен летать и, сидя на жердочке, сохранять равновесие, но склонен часами оставаться в покое. Под действием раздражения он двигается, хотя и бесцельно, как бы наудачу; так как он не способен есть предлагаемую пищу, то при отсутствии искусственного питания он может умереть с голоду. Собака после удаления коры больших полушарий способна ходить и проглатывать пищу, вложенную ей в рот, но не обнаруживает никаких проявлений страха или возбуждения. У человека иногда рождаются младенцы с неразвившейся корой больших полушарий, и, хотя они способны к осуществлению вегетативных функций дыхания и глотания, они ничему не научаются в результате опыта и не делают никаких произвольных движений. Такие дети обычно умирают вскоре после рождения.
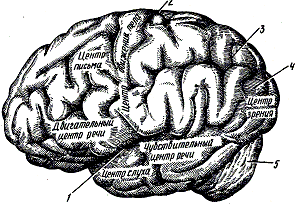
Рисунок 303. Передний мозг. 1 - сильвиева борозда; 2 - центральная борозда; З - теменно-затылочная борозда; 4 - затылочная доля; 5 - мозжечок.
В больших полушариях сосредоточено несколько больше половины всех 10 млрд. нейронов, из которых состоит нервная система человека. Большие полушария развиваются как выросты переднего конца головного мозга. У человека и других млекопитающих они растут назад, поверх остальных частей мозга и прикрывая их. Каждое полушарие содержит полость, соединенную каналом с третьим желудочком (в таламусе). Это первый и второй мозговые желудочки; в них, как и в остальных желудочках, находятся сплетения кровеносных сосудов, выделяющих цереброспинальную жидкость. Большие полушария состоят из серого и белого вещества; белое вещество, образованное из нервных волокон, расположено внутри, тогда как серое вещество, состоящее из тел нервных клеток, находится на поверхности, образуя кору больших полушарий. Глубоко в веществе больших полушарий лежат другие массы серого вещества — нервные центры, служащие промежуточными станциями на пути в кору и из коры. У низших позвоночных, у которых серого вещества мало, поверхность больших полушарий гладкая, у человека же и у других млекопитающих она покрыта извилинами. Таким образом, получаются валики, разделенные бороздами, что увеличивает пространство, занимаемое серым веществом коры. Рисунок этих извилин одинаков у всех людей независимо от их умственных способностей. «География» коры больших полушарий подвергалась тщательному изучению. Представление о том, что определенные участки головного мозга имеют особые функции, возникло давно; еще «наука» френология основывалась на предположении, что функции в мозгу локализованы определенным образом и что если человек особо одарен в какой-либо области, то определенный участок мозга должен быть увеличен и вызывает образование шишки на голове. Думали, что анализ подобных «шишек» может показать, для какой деятельности больше всего пригоден данный человек.
Экспериментальные данные позволили установить, что функции в коре в значительной степени локализованы. Путем хирургического удаления у подопытных животных отдельных участков коры оказалось возможным строго локализовать многие функции; наблюдая параличи или потерю чувствительности у людей с повреждениями или опухолями головного мозга, а затем (после смерти больного) исследуя мозг и определяя локализацию поражения, удалось составить «карту» также и человеческого мозга. Во время операций на головном мозгу хирурги раздражали небольшие участки электрическим током и наблюдали, какие мышцы при этом сокращаются; поскольку мозговые операции могут производиться под местным обезбо-ливанием, больного можно было спросить, какие ощущения он испытывает при раздражении того или иного участка. Интересно, что в самом мозгу нет нервных окончаний, воспринимающих боль; поэтому раздражение коры безболезненно. Новейший метод изучения активности головного мозга состоит в измерении и регистрации электрических потенциалов, или волн, возникающих в различных участках мозга.
По наружной стороне полушария сверху вниз проходит легко распознаваемая глубокая борозда (роландова борозда), которая отделяет двигательную зону, контролирующую скелетные мышцы, от лежащей позади нее области, ответственной за ощущение тепла, холода, прикосновения и давления при раздражении рецепторов кожи. Внутри обеих зон имеет место дальнейшая специализация участков вдоль борозды (от верхушки мозга к его боковой стороне): нейроны верхнего участка контролируют мышцы ступни, клетки последующих участков — мышцы голени, бедра, живота и т.д., а нейроны нижнего участка управляют мышцами лица. Величина корковой двигательной зоны для той или иной части тела пропорциональна не количеству мышечной ткани, а тонкости и сложности движений; особенно обширны, например, зоны, управляющие кистью руки и лицом. Аналогичное соотношение существует между частями сенсорной зоны и участками кожи, с которых они получают импульсы. Таким образом, в связях между телом и головным мозгом мы наблюдаем не только перекрещивание волокон, в результате чего правая половина мозга контролирует левую половину тела — и наоборот, но и еще одну инверсию, в результате которой самый верхний участок коры управляет самыми нижними частями тела.
В головном мозге имеются нервные центры, управляющие собственно человеческими способностями: умом, речью, памятью и т.д. (рис. 304). Эти важные функции выполняются не всем головным мозгом, общий вес которого составляет всего 1,5 кг. Сигналы, передаваемые через нервные пути, поступают только в кору головного мозга, состоящую из серого вещества. Там же локализованы и чисто человеческие функции.
Чувствительная и двигательная зоны произвольных мышц находятся соответственно в лобной и теменной долях.
Нервные центры чувств расположены в конкретных долях, и рядом с каждым из них существует архив, или центр памяти. Например, центр зрительной памяти можно сравнить с фотографическим архивом, в котором имеется карточка с изображением и названием каждого известного нам предмета.
Некоторые умственные способности локализуются в лобных долях, у других нет точного местонахождения. Мышление и речь, то есть способность облекать мысли в слова, — чисто человеческие свойства.
 Центр речинаходится в левом полушарии головного мозга, и именно в этом центре образуется понятие, выражаемое каждым словом. Другие близлежащие центры содержат «архивы» значения слов, «ищут» нужные нам слова для выражения того, что мы хотим сказать. Следующий шаг — это овеществление мысли через нервные импульсы, которые приводят в движение речеобразующие органы (устная речь) или управляют мышцами руки и кисти (письменная речь).
Центр речинаходится в левом полушарии головного мозга, и именно в этом центре образуется понятие, выражаемое каждым словом. Другие близлежащие центры содержат «архивы» значения слов, «ищут» нужные нам слова для выражения того, что мы хотим сказать. Следующий шаг — это овеществление мысли через нервные импульсы, которые приводят в движение речеобразующие органы (устная речь) или управляют мышцами руки и кисти (письменная речь).
Во время сна организм восстанавливает энергию, израсходо-
Рисунок 304. ванную в течение дня; произвольные мышцы расслабляются, а некоторые непроизвольные, такие как дыхательные, замедляют свою работу. Однако покой нервной системы лишь частичен, так как продолжается работа головного мозга.
Эта деятельность отражается в сновидениях, которые бывают всегда, хотя проснувшись, их часто не помнят. Сны — это своего рода «отдушина» нашего подсознания.
Этот механизм состоит из различных этапов, в которых фазы «медленного» сна сменяются фазами «быстрого». Именно в фазах «быстрого» сна у нас бывают сновидения: если мы спим 8 часов, то видим сны в течение четырех или пяти фаз, продолжительностью 15-20 минут каждая.
Память — одна из главных функций головного мозга. Без нее мы не могли бы ничему научиться и не извлекли бы никакой пользы из опыта. Память не локализована в какой-либо конкретной зоне коры головного мозга. То, что мы усваиваем, рассредоточивается по бесчисленным взаимосвязанным нейронам.
Полагают, что память базируется в ядре нейронов, которые не претерпевают никаких изменений, когда информация хранится в кратковременной памяти (номер телефона, урок, который мы изучаем и т.д.), но подвергаются химическим изменениям при хранении информации в долговременной памяти (пережитый опыт, воспоминания и т. д.).
Существует связь между памятью и эмоциями, так как обычно мы помним лучше то, что нам приятно или, наоборот, то, что является очень неприятным. Механизм забывания действует таким же образом: работает как защитная система, стирается то, что вызывает у нас страх или тревогу.
Головной и спинной мозг одеты тремя оболочками (рис. 305-307): твердой, паутинной и сосудистой. Твердая — наружная, соединительнотканная, выстилает внутреннюю полость черепа и позвоночного канала. Паутинная расположена под твердой — это тонкая оболочка с небольшим количеством нервов и сосудов. Сосудистая оболочка сращена с мозгом, заходит в борозды и содержит много кровеносных сосудов. Между сосудистой и паутинной оболочками образуются полости, заполненные мозговой жидкостью.
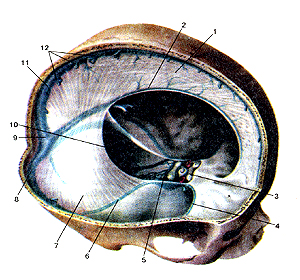
Рисунок 305. Твердая оболочка головного мозга (dura mater encephali). Синусы твердой оболочки. 1 - серп большого мозга; 2 - нижний сагиттальный синус; 3 - передний межпешеристый синус; 4 - клиновидно-теменной синус; 5 - задний межпещеристый синус; 6 -верхний каменистый синус; 7 - намет мозжечка; 8 - поперечный синус; 9 - синусный сток; 10 -сигмовидный синус; 11 - верхний сагиттальный синус; 12 - устья верхних мозговых вен.
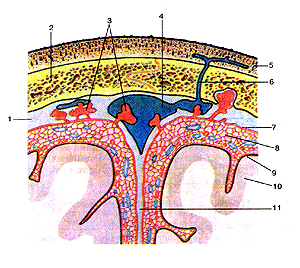
Рисунок 306. Оболочки головного мозга на поперечном (фронтальном) paзpeзе. Взаимо-расположение оболочек и верхнего сагиттального синуса со сводом черепа и поверхностью головного мозга. 1 - твердая оболочка головного мозга; 2 - свод черепа; 3 - грануляции паутинной оболочки; 4 - верхний сагиттальный синус; 5 - кожа; 6 - эмиссарная вена; 7 -паутинная оболочка головного мозга; 8 - подпаутинное пространство; 9 - мягкая оболочка головного мозга; 10 - головной мозг; 11 - серп большого мозга.
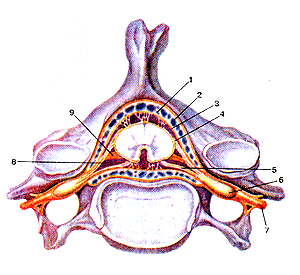
Рисунок 307. Оболочки спинного мозга (meninges medullae spinalis) в позвоночном канале. По-перечный разрез на уровне межпозвоночного диска. I - твёрдая оболочка спинного мозга; 2 -эпидуральное пространство; 3 - паутинная оболочка; 4 - задний корешок спинномозговой нерва; 5 - передний корешок; 6 - спинномозговой узел; 7 - спинномозговой нерв; 8 - подпаутин-ное (субарахноидальное) пространство; 9 - зубчатая связка.
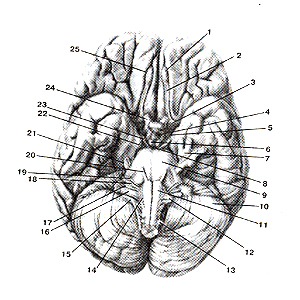
Рисунок 308. Нижняя поверхность (основание) головного мозга и места выхода корешков черепных нервов. I - обонятельная луковица; 2 - обонятельный тракт; 3 - переднее продырявленное вещество; 4 - серый бугор; 5 - зрительный тракт; 6 - сосцевидные тела; 7 -тройничный узел; 8 - заднее продырявленное вещество; 9 - мост; 10 - мозжечок; 11 - пирами-да; 12 - олива; 13 - спииномозровой нерв; 14 - подъязычный нерв; 15 - добавочный нерв; 16 -блуждающий нерв; 17 - языкоглоточный нерв; 18 - преддверно-улитковый нерв; 19 - лицевой нерв; 20 - отводящий нерв; 21 - тройничный нерв; 22 - блоковый нерв; 23 - глазодвигательный нерв; 24 - зрительный нерв; 25 - обонятельная борозда.
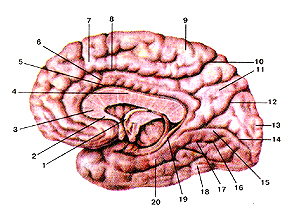
Рисунок 309. Медиальная и нижняя поверхности полушария большого мозга. 1 - свод; 2 -клюв мозолистого тела; 3 - колено мозолистого тела; 4 - ствол мозолистого тела; 5 - борозда мозолистого тела; 6 - поясная извилина; 7 - верхняя лобная извилина; 8 - подтеменная борозда; 9 - парацентральная долька; 10 - поясная борозда; 11 - предклинье; 12 - теменно-затылочная борозда; 13 - клин; 14 - шпорная борозда; 15 - язычная извилина; 16 - медиальная затылочно-височная извилина; 17 - затылочно-височная борозда; 18 - латеральная затылочно-височная извилина; 19 - борозда гиппокампа; 20 - парагиппокампальная извилина.
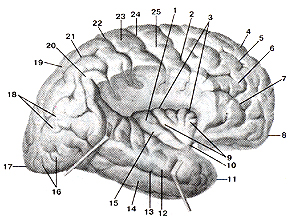
Рисунок 310. Островок (insula). Островковая доля. Вид с латеральной стороны. Часть те-менной и лобной долей удалена. Височная доля оттянута книзу. 1 - островок; 2 - пред-центральная борозда; 3 - круговая борозда островка; 4 - верхняя лобная извилина; 5 - верхняя лобная борозда; 6 - средняя лобная извилина; 7 - нижняя лобная борозда; 8 - лобный (передний) полюс; 9 - короткие извилины островка; 10 - порог островка; 11 - височный полюс; 12 - верхняя височная извилина; 13 - верхняя височная борозда; 14 - средняя височная извилина; 15 - длинная извилина островка; 16 - латеральные затылочные извилины; 17 - затылочный (задний) полюс; 18 - угловая извилина; 19 - верхняя теменная долька; 20 - надкраевая извилина; 21 - внут-ритеменная борозда; 22 - постцентральная борозда; 23 - постцентральная извилина; 24 -центральная борозда; 25 - предцентральная извилина.
Продолговатый мозг (рис. 311) — это самый задний отдел головного мозга, лежащий 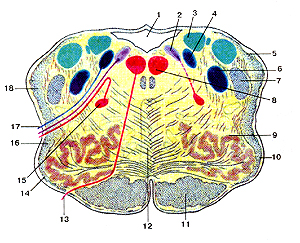
Рисунок 311. Продолговатый мозг (medulla oblongata). Поперечный разрез на уровне олив. I -четвертый желудочек; 2 - дорсальное ядро блуждающего нерва; 3 - ядро вестибулярного нерва; 4 - ядро одиночного пути; 5 - задний (дорсальный) спинно-мозжечковый путь; 6 -спинномозговое ядро тройничного нерва; 7 - спинно-мозговой путь тройничного нерва; 8 - ядро подъязычного нерва; 9 - оливное ядро; 10 - олива; 11 - корково-спинномозговой путь (пирамидный); 12 - медиальная петля; 13 - подъязычный нерв; 14 - передние наружные дуговые волокна; 15 - двойное ядро; 16 - спинно-таламический и спинно-покрышечный пути; 17 -блуждающий нерв; 18 - вентральпый (передний) спинно-мозжечковый путь.
непосредственно спереди от спинного мозга. Здесь центральный канал спинного мозга расширяется и образует большую полость, называемую четвертым мозговым желудочком (три остальных желудочка находятся выше, в передних отделах головного мозга). Крыша четвертого желудочка тонка и содержит сплетение кровеносных сосудов, выделяющих часть цереброспинальной жидкости; остальную часть этой жидкости выделяют аналогичные сплетения, находящиеся в других желудочках. В крыше четвертого желудочка имеется три крошечных отверстия, через которые цереброспинальная жидкость выходит в пространства между мозговыми оболочками. Стенки продолговатого мозга толстые и состоят главным образом из нервных путей, идущих к высшим отделам головного мозга. В сером веществе продолговатого мозга залегают жизненно важные центры, регулирующие сердечную деятель-ность, дыхание, глотание, осуществляющие защитные рефлексы (чихание, кашель, рвота, слезоотделение), секрецию слюны, желудочного и поджелудочного сока и др. Повреждение продолговатого мозга может быть причиной смерти вследствие прекращения сердечной деятельности и дыхания.
Над продолговатым мозгом расположен мозжечок (рис. 312, 313), состоящий из средней
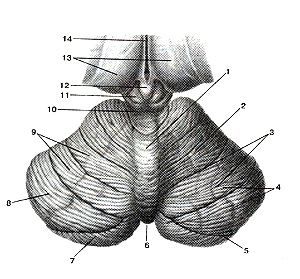
Рисунок 312. Мозжечок (cerebellum). Вид сверху. 1 - червь мозжечка; 2 - полушарие мозжечка; 3 - щели (борозды) мозжечка; 4 - листки мозжечка; 5 - горизонтальная щель; 6 -задняя вырезка мозжечка; 7 - нижняя полулунная долька; 8 - верхняя полулунная долька; 9 -четырехугольная долька; 10 - нижние холмики крыши среднего мозга; 11 - верхний холмик; 12 -эпифиз; 13 - таламусы; 14 - третий желудочек.
части и двух боковых полушарий, по форме напоминающих сосновые шишки. Серый поверхностный слой мозжечка состоит из тел нервных клеток, а глубже находится масса белой ткани, образованной волокнами, связывающими мозжечок с продолговатым мозгом и с высшими отделами мозга. Весь мозжечок представлен двумя полушариями, средней частью - червем и тремя парами ножек, образованных нервными волокнами, с помощью которых он связан с другими отделами головного мозга. Основная функция мозжечка — безусловно-рефлекторная координация движений, определяющая их четкость, плавность и сохранение равновесия тела, а также поддержание тонуса мышц. Через спинной мозг по проводящим путям импульсы от мозжечка поступают к мышцам. Контролирует деятельность мозжечка кора больших полушарий.
|
|
|
|
|
Дата добавления: 2014-11-07; Просмотров: 1350; Нарушение авторских прав?; Мы поможем в написании вашей работы!