КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вопросы. 1. Строение и физиология наружного и среднего уха
|
|
|
|
1. Строение и физиология наружного и среднего уха.
2. Строение и физиология внутреннего уха.
3. Нейроанатомическая схема слуховой системы человека.
4. Частотно-пороговые характеристики нейронов слуховой системы.
Литература
Гильберт С. Слух. Введение в психологическую и физиологическую акустику. М.: Медицина, 1984.
НЕЙРОФИЗИОЛОГИЯ СОМАТОСЕНСОРНОЙ СИСТЕМЫ
Спинной мозг является главным коллектором (собирателем) сенсорных импульсов с поверхности тела. Каждый дорсальный корешок данного сегмента спинного мозга собирает информацию с поверхности тела, которая называется дерматомом. В приложении 8 показана схема таких дерматомов тела человека. Центральная веточка дорсального корешка спинного мозга содержит аксоны разного диаметра, которые проводят импульсацию от различных рецепторов. Толстые миелинизированные аксоны сообщают информацию о прикосновении и от суставных рецепторов. Тонкие нервные волокна, среди которых много немиелинизированных, проводят болевую и температурную чувствительность. После прихода нервных волокон в серое вещество дорсального рога они многократно ветвятся и образуют три типа окончаний: сегментарные (т.е. в пределах одного сегмента спинного мозга), восходящие (часть из них участвует в формировании восходящих путей спинного мозга) и нисходящие (участвуют в осуществлении межсегментарных рефлексов).
Кожа как сенсорная система. Кожа отграничивает организм от внешней среды; она содержит многообразные рецепторы, которые сигнализируют организму о контактах с внешним миром. Кожа состоит из трех слоев: эпидермиса, дермы (собственно кожи) и подкожной клетчатки (рис. 6.20). Под эпидермисом в слое соединительной ткани находится густая сеть нервных волокон с сенсорными окончаниями в эпидермисе и дерме. В дерме и подкожной клетчатке эти волокна собираются в пучки, а затем в крупные нервы, которые направляются в составе задних корешков в спинной мозг.
Рецепторные структуры кожи бывают двух видов: свободные и инкапсулированные окончания нервных волокон. Свободные окончания часто контактируют с клеткой-сателлитом эпидермального происхождения. Однако участвует ли эта клетка в реакции рецептора,- неизвестно. Самым простым типом рецептора кожи является свободный (не покрытый миелином) кончик тонкого афферентного волокна. Свободные нервные окончания находятся между клетками эпидермиса. По-видимому, значительная часть свободных окончаний представляет собой ноцицептивные рецепторы (рецепторы повреждения, болевые). Другие свободные нервные окончания связаны с ощущениями зуда, щекотки; ими же, по-видимому, воспринимается температура (терморецепторы). Некоторые из немиелинизированных волокон являются эфферентными постганглионарными симпатическими волокнами, которые иннервируют гладкие мышцы сосудистой стенки и волосяные фолликулы. Распространенным типом рецепторов являются также свободные нервные окончания, которые окружают волосяные фолликулы. Эти рецепторы возбуждаются при смещении волоса или его подергивании. В области волосяной фолликулы имеются также инкапсулированные окончания толстых миелинизированных волокон, которые, по-видимому, возбуждаются при сгибании волоса.
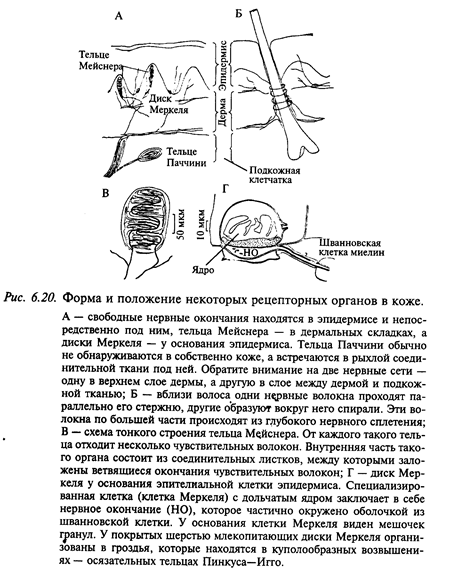
Инкапсулированные окончания составляют важный класс рецепторов кожи. Роль капсулы из соединительной ткани весьма значительна в формировании реакции рецептора. В настоящее время выделяют следующие инкапсулированные окончания кожи: диски Меркеля, тельца Мейснера, тельца Паччини, окончания Руффини. При изучении свойств кожных рецепторов трудно бывает сопоставить их физиологические характеристики с данными гистологии. Это связано с тем, что при регистрации биоэлектрической активности от афферентного волокна достаточно просто и надежно можно определить рецептивное поле данного афферентного волокна, но в этом поле могут находиться сотни рецепторов, которые передают информацию по другим афферентным волокнам. Поэтому очень трудно сопоставить тип реакции афферента с морфологическими типами рецепторов в данном участке кожи. Другими словами, можно дать физиологическую классификацию по реакциям на стимулы и распределить наблюдаемые под микроскопом рецепторы по морфологическим классам. Однако сопоставить функциональную и анатомическую классификации удается далеко не всегда. На рис. 6.20 схематически показаны те виды рецепторов, которые можно обнаружить у млекопитающих в коже, покрытой волосами (А) и не покрытой волосами (Б).
Рецепторы кожи можно разделить на быстро адаптирующиеся, или фазические (например, тельца Паччини); медленно адаптирующиеся, или тонические (например, диски Меркеля, окончания Руффини), и имеющие среднюю скорость адаптации, или фазно-тонические (например, рецепторы волосяных луковиц, тельца Мейснера).
Проприорецепторы. Известно, что млекопитающие могут достаточно точно оценивать положение конечностей, головы и туловища в пространстве. Кроме этого, достаточно точно оцениваются величина сопротивления каждому движению, углы суставов, скорость их изменения и пр. Такая оценка становится возможной благодаря рецепторам, которые находятся в мышцах, сухожилиях, суставах и называются проприорецепторами. Выше были описаны физиологические особенности двух проприорецепторов - мышечных веретен и сухожильных рецепторов. Сигналы от рецепторов кожи, мышц, суставов, сухожилий и внутренних органов поступают через задние корешки в спинной мозг. Здесь эта информация обрабатывается и отсюда передается в верхние (супраспинальные) этажи ЦНС: ядра вентробазального таламуса и далее в кору больших полушарий.
Механорецепция объединяет четыре качества: ощущение давления, прикосновения, вибрации и щекотания. Изучают механорецепцию одним из классических методов: прикосновение к коже волосками, или щетинками, позволило установить, что даже легкое надавливание (в диапазоне 0,1-0,5 г, что приводит к вдавливанию кожи всего на 0,1-0,01 мм) вызывает ощущение прикосновения или давления. Но эти ощущения возникают только при надавливании в определенных точках, например кожи ладони. Эти точки называются тактильными точками. Участки кожи с большой плотностью тактильных точек находятся на кончиках пальцев и на губах. Плотность тактильных точек на животе, спине и плечах очень низкая.
Используя ножки специального циркуля (с тупыми концами, чтобы избежать болевого раздражения), можно определять пороги пространственного различения, т.е. минимальные расстояния между двумя одновременными тактильными стимулами, при которых их удается воспринять как раздельные. Исследования показали, что у взрослого человека одновременные пространственные пороги кончиков пальцев и губ особенно низки (1-3 мм), тогда как на спине, плечах и бедрах они существенно выше (50-100 мм), что согласуется с плотностью тактильных точек на этих частях тела человека.
Чувствительность к вибрации часто изучают при обследовании больных. Целью такого обследования чаще всего является нахождение порога субъективного ощущения вибрации. Наибольшая чувствительность к вибрации у человека находится в диапазоне 150 - 300 Гц. Минимальная (пороговая) амплитуда вибрации в этом частотном диапазоне составляет около 1 мкм.
Боль радикально отличается от других ощущений тем, что передает в нервный центр сигнал тревоги. Модальность «боль» объединяет два типа соматических болей и висцеральные боли.
Если соматическая боль имеет кожное происхождение, ее называют поверхностной болью. Если источник боли находится в мышцах, суставах или соединительной ткани, она называется глубокой болью. Таким образом, поверхностные и глубокие боли это два подкласса соматической боли. При реальном ощущении боли выделяют начальную боль, ощущаемую человеком как вспышку боли, которая исчезает с прекращением стимуляции. За этой резкой и легко локализуемой начальной болью часто следует отставленная боль, имеющая ноющий тупой характер, она начинает ощущаться через 0,5-1 с.
Болевые рецепторы в коже расположены в виде отдельных точек (дискретно). Поэтому в тех участках кожи, где их плотность невелика, всегда можно найти точки, лишенные болевых рецепторов. Специальные исследования показали, что кожа человека содержит значительно больше болевых точек, чем тактильных (соотношение 9:1). Температурные рецепторы (точки холода и тепла) встречаются в коже еще реже, чем тактильные точки, поэтому их соотношение с болевыми точками примерно 10:1.
Висцеральная боль (внутренних органов) также по своему характеру похожа на глубокую боль - имеет тупой диффузный характер. Примечательно, что внутренние органы (например, при вскрытии брюшной полости под местной анестезией) при сжимании и даже разрезах не вызывают ощущения боли, если только не задевать париетальную брюшину и корень брыжейки. Однако растяжение полых органов вызывает резкое ощущение боли. Кроме того, болью сопровождаются спазмы или сильные сокращения гладких мышц, особенно если этому сопутствует нарушение кровообращения (ишемия).
Болевые сигналы от внутренних органов часто приводят к болевым ощущениям не только в этих органах, но и распространяются на соответствующие дерматомы. Это так называемая отраженная боль. Примером такой боли, может быть, боль в сердце, которая ощущается в груди и узкой полоске на медиальной поверхности руки.
НЕЙРОФИЗИОЛОГИЯ СЕНСОРНЫХ ПУТЕЙ СПИННОГО МОЗГА
1. Дорсальные (задние) столбы спинного мозга представляют собой мощную систему волокон различного происхождения (рис. 6.21). Задние столбы у наземных млекопитающих в эволюции формируются в связи с развитием конечностей, когда возникает необходимость в тонком анализе информации от суставов и кожи, без которого невозможны ходьба, бег, прыжки, удержание равновесия и положения тела в пространстве. Прогрессивное развитие задних столбов в эволюции позвоночных животных тесно связано с развитием соматосенсорной коры больших полушарий.
Волокна задних корешков, войдя в спинной мозг, делятся на две веточки: длинную (восходящую) и короткую (нисходящую). Ядер задних столбов (нежного и клиновидного) в продолговатом мозге достигают лишь около четверти миелинизированных волокон, остальные аксоны оканчиваются на интернейронах спинного мозга.
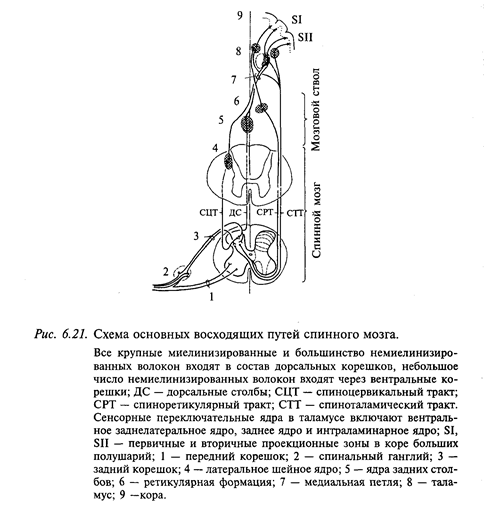
Аксоны нейронов ядер задних столбов (нежного и клиновидного) перекрещиваются и образуют медиальные лемниски, которые направляются к ядрам вентробазального таламуса. Волокна, составляющие задние столбы в спинном мозге, постепенно истончаются, так как по ходу отдают большое число коллатералей к интернейронам спинного мозга, поэтому скорость проведения импульсов снижается с 90-120 м/с - в месте вхождения в спинной мозг до 20-40 м/с на уровне шейных верхних сегментов спинного мозга.
В системе волокон задних столбов проходят афференты от многих типов рецепторов, в частности у животных - от рецепторов волосяных луковиц, суставов, подушечек лап, основания когтей и других образований. Мышечные афференты от конечностей также восходят в составе задних столбов и заканчиваются на нейронах ядер продолговатого мозга, а также на нейронах дорсального рога шейных верхних сегментов спинного мозга. Аксоны этих нейронов 2-го порядка перекрещиваются и присоединяются к волокнам медиального лемниска, которые, переключаясь через ядра вентробазального комплекса, проецируются на кору больших полушарий на границе сенсорной и моторной зон (поле За, по Бродману). Значительная часть афферентных волокон от мышечных и сухожильных рецепторов не входит в состав задних столбов. Эти волокна, начиная с поясничных сегментов спинного мозга и выше, переключаются на нейронах кларковых столбов, аксоны которых образуют мощные сенсорные тракты к мозжечку.
Суставные сумки и поверхности суставов имеют многообразные рецепторы, которые объединяются под общим названием суставные рецепторы. Согласно современным данным, в суставном нерве выделяют два типа ответов: быстро адаптирующиеся и медленно адаптирующиеся, последние более многочисленны. Соответственно этому выделяют два типа рецепторов. Медленно адаптирующиеся рецепторы, являясь наиболее типичными суставными рецепторами, сообщают о положении костей, образующих данный сустав. В суставных нервах обнаруживаются волокна, в которых разряды возникают как при сгибании, так и при разгибании сустава. Обнаружены также волокна, разряд которых в зависимости от сгибания или разгибания сустава, учащается или урежается (дирекционно-чувствительные ответы). Одно и то же движение сустава может вызвать урежение импульсации в одном афферентном волокне и увеличение ее в другом волокне. Информация от суставных рецепторов в супраспинальные центры проводится по системе волокон задних столбов.
Волокна задних столбов проводят также афферентные сигналы от внутренних органов. Импульсы от тазового нерва достигают надсегментарных структур не только по задним канатикам (задним столбам), но также и по вентролатеральным канатикам, в составе спиноталамических трактов спинного мозга. Скорость проведения импульсов от внутренних органов в задних столбах, например, кошки составляет 35-75 м/с, в вентральных-21-38 м/с и латеральных-17-32 м/с.
2. Спиноцервикальный тракт образован аксонами нейронов, тела которых находятся в основании серого вещества дорсального рога спинного мозга (см. рис. 6.21). Далее волокна тракта проходят ипсилатерально в составе латерального канатика и оканчиваются на нейронах шейного латерального ядра, локализованного в верхних сегментах шейного отдела серого вещества задних рогов спинного мозга. На этом же уровне аксоны нейронов этого ядра перекрещиваются и в составе специальной порции медиального лемниска направляются в вентробазальный комплекс таламических ядер.
Известно, что шейное латеральное ядро у человека представлено рассеянными клетками в шейных сегментах спинного мозга. У кошки, собаки, низших обезьян это ядро развито очень хорошо. На нейронах спиноцервикального тракта конвергирует возбуждение от низкопороговых кожных рецепторов, в том числе от рецепторов волосяных луковиц. Ряд нейронов возбуждается при пощипывании или давлении на кожу; размер их рецептивных полей может быть большим и занимать даже всю ипсилатеральную половину тела. Несмотря на большее количество переключений (спинальный уровень, шейное латеральное ядро, вентробазальный таламус, кора больших полушарий), возбуждение по спиноцервикальному тракту прибывает в кору больших полушарий на 2-5 мс быстрее, чем по волокнам дорсальных столбов. Это объясняется тем, что скорость проведения по волокнам задних столбов меньше, чем по волокнам спиноцервикального тракта.
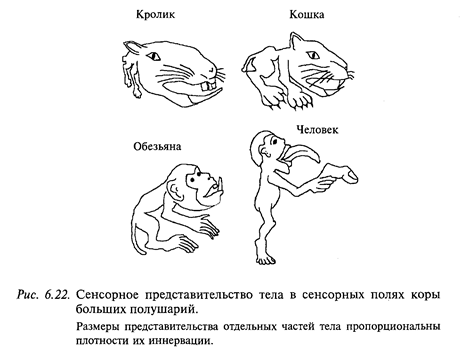
У млекопитающих (в том числе и у человека) по дорсальным столбам и спиноцервикальному тракту (у человека преимущественно по дорсальным столбам) в постцентральной коре больших полушарий организуется проекция всего тела (рис. 6.22). При этом соблюдается следующий принцип: чем выше плотность рецепторов в коже, покрывающей соответствующую часть тела, тем большей площадью эта проекция представлена в коре больших полушарий. Обратите внимание на то, что руки, особенно большой палец, оральная область, в том числе язык, голосовые связки и пр. представлены в коре мозга человека особенно подробно.
3. Спиноталамический тракт составляют самые тонкие миелинизированные афференты (диаметром до 6 мкм), а также немиелинизированные волокна задних корешков. Они входят в серое вещество спинного мозга и заканчиваются на нейронах основания серого вещества заднего рога. В сером веществе спинного мозга аксоны части этих нейронов перекрещиваются, а другая часть идет ипсилатерально, образуя спиноталамический тракт. Таким образом, спиноталамический тракт проецируется в ядра таламуса (вентральное заднелатеральное, парафасцикулярное и центральное латеральное ядра) билатерально. Часть нервных волокон тракта переключается на нейронах мозгового ствола, аксоны части этих нейронов переходят на контралатеральную сторону и также направляются в таламус.
По волокнам спиноталамического тракта передается информация о боли, давлении и температуре. Нейроны этого тракта имеют обширные кожные рецептивные поля, располагающиеся у некоторых нейронов билатерально, покрывая обе задние или обе передние конечности, а иногда даже всю поверхность тела. Нейроны, активируемые при раздражении одной конечности, как правило, тормозятся при стимуляции другой, тогда как нейроны с ограниченными кожными рецептивными полями часто тормозятся с окружающей кожной поверхности. В настоящее время считают, что этот тракт проводит генерализованные ощущения и по нему передается информация только о качестве раздражителя и его грубой локализации.
Таким образом, в таламус проецируются две системы: задние столбы (у ряда видов млекопитающих также спиноцервикальная система) и спиноталамический тракт. Эти две системы различаются функционально: первая проводит информацию о месте и модальности раздражителя (протопатическая чувствительность), тогда как вторая обеспечивает генерализованные формы ощущения (эпикритическая чувствительность).
ФИЗИОЛОГИЯ ТРОЙНИЧНОГО НЕРВА
Ветви тройничного нерва осуществляют иннервацию различных тканей головы. Большинство рецепторов этой системы возбуждаются только при непосредственном контакте с раздражителем. Примером высокой специализации может служить фациальная ямка некоторых змей, с помощью которой эти животные эффективно локализуют на расстоянии источник теплового излучения. У млекопитающих телерецепция такого типа отсутствует, но у них хорошо развиты вибриссы, с помощью которых животные активно исследуют окружающую среду. Самой высокой сложности тригеминальная система достигает у человека в связи с ее участием в речевой функции.
Все многочисленные ветви тройничного нерва объединяются в три подразделения: глазничное, максиллярное и мандибулярное (рис. 6.23). У входа в головной мозг они образуют крупный полулунный (гассеров) ганглий, образованный телами псевдоуниполярных нейронов. Зрительная ветвь иннервирует кожу лба и носа, роговицу, конъюнктиву и внутриглазные структуры, слизистую параназальных синусов, верхней, нижней и передней частей перегородки носа и латеральной стенки носовой полости, слезный проток. Верхнечелюстная ветвь иннервирует область кожи верхней челюсти, слизистую максиллярного синуса, верхние зубы и десны, слизистую задней части перегородки носа и нижней части носовой полости, верхние зубы и десны, твердое небо и миндалины. Нижнечелюстная ветвь иннервирует область кожи нижней челюсти, слизистую щек, дно носовой полости, язык, проприорецепторы мышц нижней челюсти, нижние зубы и десны, жевательные мышцы. Все подразделения тройничного нерва иннервируют мозговые оболочки и сосуды мозга.
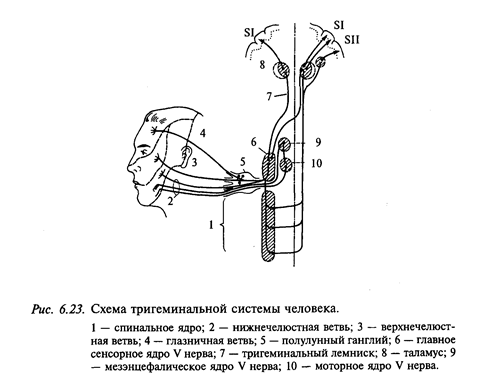
Некоторая часть первичных афферентов зрительной и верхнечелюстной ветвей, а также значительное количество волокон мандибулярной ветви не имеют клеточных тел в гассеровом ганглии. Тела этих афферентных нейронов находятся в тройничном мезэнцефалическом ядре, которое представляет собой узкую полоску псевдоуниполярных нейронов, протянувшуюся вдоль всего среднего мозга. С филогенетической точки зрения тройничное мезэнцефалическое ядро, как и гассеров ганглий, является гомологом спинномозговых ганглиев. Аксоны нейронов этого ядра проходят с моторными ветвями тройничного нерва к жевательным мышцам и осуществляют афферентную иннервацию проприорецепторов (мышечных веретен и сухожильных рецепторов). Они также иннерви-руют рецепторы зубов, языка, сустава нижней челюсти; при этом большая часть их направляется ипсилатерально, но некоторые отсылают свои аксоны на контралатеральную сторону. Часть волокон мезэнцефалического ядра заканчивается в моторном тройничном ядре, образуя двухнейронную дугу жевательных рефлексов, но некоторая часть спускается к ядрам спинальных тригеминальных ядер, расположенных на уровне продолговатого мозга.
Центральные отростки нейронов мезэнцефалического тригеминального ядра объединяются в латеральный продольный пучок, который, таким образом, является афферентной частью рефлекторных систем, осуществляющих контроль кусания, жевания, глотания, участвует в сокращении наружных глазных мышц и мимической мускулатуры. Афференты сенсорного корешка тройничного нерва в мозге делятся на толстые нисходящие и тонкие восходящие волокна. Восходящая часть волокон направляется к главному сенсорному ядру (на уровне моста), а более толстые нисходящие волокна образуют нисходящий спинальный тройничный тракт, который спускается вниз и достигает уровня первых шейных сегментов, синаптически контактируя с ядрами тройничного тракта. В составе тройничного тракта, кроме первичных афферентов, имеется значительное количество аксонов нейронов 2-го порядка.
В составе волокон тройничного тракта имеются волокна лицевого, языкоглоточного и блуждающего нервов. В настоящее время различают два компонента тригеминального комплекса: главное сенсорное ядро (локализовано в мосте) и сенсорные ядра спинотригеминального тракта (локализованы в продолговатом мозге и верхних шейных сегментах). Филогенетические данные и характер центральных связей тройничных сенсорных ядер позволяют рассматривать главное сенсорное ядро как гомолог ядер дорсальных столбов спинного мозга, тогда как ядра тригеминального тракта как гомолог серого вещества дорсальных рогов спинного мозга.
В составе тройничных нервов идут также волокна вегетативной нервной системы. Парасимпатические волокна берут начало от верхнего и нижнего слюнных ядер продолговатого мозга. Симпатические волокна являются аксонами нейронов боковых рогов спинного мозга. Вегетативные волокна, входящие в состав ветвей тройничного нерва, регулируют секрецию желез слизистой оболочки рта и носа.
Функциональные характеристики афферентных волокон тройничного нерва. Афферентные волокна тройничного нерва проводят афферентную импульсацию разной модальности - от механорецепторов, терморецепторов и болевых рецепторов. Окончания тройничного нерва образуют два типа терморецепторов: специфические терморецепторы реагируют только на изменение температуры, неспецифические - как на температурные, так и на механические воздействия. Терморецепторы у большинства млекопитающих локализованы в структурах ротовой полости и кожи носа. Пульпа зубов содержит только болевые рецепторы, которые иннервируются толстыми волокнами.
Механорецепторы, от которых идет импульсация по волокнам тройничного нерва, тонко специализированы, что обеспечивает точность оценки механического стимула в области морды животного. Особенно тонкий анализ механических стимулов у млекопитающих осуществляется рецепторами волос. У них выделяют три типа волос: вибриссы (сенсорные волосы), остевые волосы (тилотрихи) и волосы шерсти. Каждому типу волос соответствуют особенности строения волосяных луковиц. Особенно велико значение вибрисс. У крысы, например, одну луковицу вибриссы иннервируют 100-150 нейронов гассерова ганглия, при этом отдельные нейроны передают в мозг информацию о таких признаках, как исходное положение вибриссы, направление, скорость и амплитуда ее отклонения и т. д. Афферентные волокна, передающие импульсы от механорецепторов в области морды, имеют очень маленькие рецептивные поля по сравнению с афферентными волокнами, иннервирующими поверхность лап или туловища. Например, рецептивные поля на морде кошки имеют размер 7 - 15 мм2, а вибрисс - 1 мм2. На гладкой коже носа у кошки, собаки и обезьяны размер рецептивного поля - тоже порядка 1 мм2. Рецепторы растяжения наружных мышц глаза у млекопитающих иннервируются тонкими аксонами нейронов, тела которых находятся в мезэнцефалическом ядре (кошка, человек), но у некоторых животных (овца, свинья) - в гассеровом ганглии.
Проекция первичных афферентов в ядра тройничного нерва исключительно ипсилатеральная и организована в виде «листков», которые идут в рострокаудальном направлении, включая по нескольку ядер комплекса. Нейроны, получающие сигналы из полости рта, локализованы в более медиальных областях тройничных сенсорных ядер, тогда как периоральные структуры и кожа головы представлены в них латерально. У млекопитающих самую обширную группу в тройничных сенсорных ядрах образуют нейроны, связанные с тактильными рецепторами, особенно те, которые возбуждаются при сгибании вибрисс; эти нейроны занимают в ядрах дорсолатеральную часть. У опоссума, например, нейроны, связанные с тактильными рецепторами вибрисс и гладкой кожи носа, занимают 4/5 объема тройничных сенсорных ядер. Центральную часть тройничных сенсорных ядер занимает столб клеток, на которые проецируются рецепторы зубов. Она простирается от центральной области главного сенсорного ядра до рострального ядра спинального тригеминального тракта. Нейроны, связанные со специфическими терморецепторами языка, локализуются в каудальном ядре спинального тригеминального тракта. Нейроны, которые активируются от неспецифических терморецепторов кожи, обнаружены как в каудальном, так и в оральном ядрах спинального тригеминального тракта. В главном сенсорном ядре, в оральном и каудальном ядрах преобладают нейроны с локальными рецептивными полями, тогда как нейроны интерполярного ядра имеют преимущественно обширные рецептивные поля. В дорсальной части орального ядра обнаружено большое количество нейронов, на которых заканчиваются афференты от периоральной области и вибрисс.
Тройничное моторное ядро, локализованное вентральнее сенсорного ядра, состоит из большого дорсального подразделения, образованного редко расположенными крупными нейронами с хорошо развитыми дендритами. Вентральная часть ядра образована более мелкими, но более тесно расположенными нейронами. Нейроны этого ядра – мотонейроны - осуществляют эфферентную иннервацию мышечных волокон жевательных мышц. В ядре существует соматотопия: мышцы верхней части головы (жевательная и височная) иннервируются мотонейронами, расположенными в вентральном подразделении ядра, тогда как мышцы нижней части головы (наружная и внутренняя крыловидная мышцы) - нейронами дорсальных отделов ядра. Полная дифференцировка ядер тригеминальной системы заканчивается только у млекопитающих.
Связи тригеминальной системы с таламусом и мозжечком. Почти все нейроны главного тригеминального сенсорного ядра посылают свои аксоны к таламусу (см. рис. 6.23). Нейроны, аксоны которых формируют билатеральные лемнисковые тракты, составляют 2/3 вентральной части главного сенсорного ядра. Ипсилатеральный тригеминальный тракт образован аксонами нейронов, тела которых локализованы в дорсальной части ядра.
От ядер спинального тройничного тракта берут начало два тригеминоталамических пути -вентральный и дорсальный. Вентральный тригеминальный тракт начинается от каудального ядра, затем переходит на контралатеральную сторону и заканчивается в заднемедиальном вентральном ядре и частично в интраламинарных ядрах таламуса. Дорсальный тригеминальный тракт начинается от нейронов каудального ядра. Это билатеральный тракт, но с преобладанием контралатеральной компоненты. Достигнув таламуса, волокна тракта оканчиваются в заднемедиальном вентральном и интраламинарных ядрах таламуса. Мозжечок получает проекции от нейронов тригеминальных ядер. Основным источником этих волокон являются ядра спинального тригеминального тракта. Их аксоны оканчиваются преимущественно рострально в коре червя мозжечка. Таким образом, тригеминоталамические пути распределены в структурах как специфического, так и неспецифического отделов таламокортикальной системы. Нейроны главного сенсорного тригеминального ядра не имеют проекций на ретикулярную формацию, тогда как ядра спинального тригеминального тракта образуют как ипси-, так и контралатеральные проекции на парамедианное и гигантоклеточное ядра ретикулярной формации.
Таламический и корковый уровни анализа кожно-кинестетической информации. Ядра вентробазального таламуса переключают информацию о прикосновении, давлении, движении в суставах. Обработка этой информации на уровне специфических таламических ядер организована по принципу соматотопии и модальности. В отличие от этого для нейронов интраламинарных ядер, заднего и ретикулярного таламических ядер, к которым подходят волокна спиноталамических трактов, характерна обширная конвергенция ряда модальностей, включая болевую.
Соматосенсорная кора имеет два подразделения: SI (первичная соматосенсорная область) и SII (вторичная соматосенсорная область). SI у приматов занимает переднюю часть теменной доли, включая дно и заднюю стенку центральной борозды. Другая, менее изученная, область - SII - лежит на передней стенке сильвиевой извилины. Кроме этого, соматосенсорные сигналы достигают нейронов моторной прецентральной коры (поля 4, 6) и поле 5 (ростральная теменная кора). Области SI и SII тесно связаны между собой. Поле 5 получает основной вход от латерального заднего таламического ядра, а также пространственно организованный вход от полей 1 и 2. Известно также, что симметричные зоны коры, включая области SI и SII, связаны между собой через мозолистое тело. В области SI коры выделяют несколько зон, соответствующих полям За, Зb, 1 и 2 (по Бродману). Нейроны поля За получают информацию от мышечных и суставных рецепторов, нейроны поля 2 - от глубоких структур (возбуждаются при сдавливании кожи и практически не реагируют на легкое прикосновение). В отличие от этого нейроны поля Зb реагируют на легкое прикосновение к коже, сгибание отдельных волосков. Нейроны поля 1 реагируют как на кожную стимуляцию, так и на стимуляцию глубоких структур.
Характерным для сенсорных и моторных полей коры является организация нейронов, участвующих в обработке однородной информации, в вертикальные колонки. Например, нейроны одной колонки области SI реагируют на стимулы одной модальности.
Эффекты разрушения сенсорных полей коры. Избирательное разрушение поля 3 приводит к тому, что, например, обезьяна на ощупь не способна отличать мягкий предмет от твердого, шероховатую поверхность от гладкой, выпуклую от вогнутой. Разрушение поля 2 нарушает способность различать форму у предметов. Например, невозможно было обучить обезьяну отличать при ощупывании квадрат от ромба. Разрушение поля 1 вызывает достаточно легкий дефицит в дискриминации рельефа поверхности при ощупывании. Было сделано заключение, что повреждение SI (в состав которой входят указанные поля) приводит не только к дефициту пространственного различения (например, определение величины углов, характера поверхности- вогнутая или выпуклая и др.), но и к дискриминации временных событий. Например, обезьяна была не способна различать осцилляции различной частоты, приложенные к поверхности кожи.
Основные результаты экспериментальных исследований были подтверждены клиническими наблюдениями больных с поврежденной постцентральной областью коры. У таких больных нарушалась дискриминация размера и формы предметов. Например, больные не могли отличить на ощупь овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Как у человека, так и у обезьяны при повреждении постцентральной коры страдает кинестезия, что проявляется в резком ухудшении способности оценивать положение конечности при пассивном или активном ее перемещении. Страдает также оценка силы, приложенной к руке.Ощущение боли и температуры при этих повреждениях коры сохраняется.
При поражении вторичных отделов постцентральной коры у больных возникают своеобразные явления «афферентной апраксии». Они проявляются в том, что больной не способен выполнять тонкие дифференцированные движения. При поражении аналогичных зон левого (доминантного) полушария с распространением на нижние отделы (область представительства лица, губ и языка) кинестетическая апраксия может проявляться в нарушении движений речевого аппарата (афферентная моторная афазия). Нарушения проявляются в невозможности найти положение губ и языка, необходимые для артикуляции соответствующих звуков. Больные смешивают близкие артикулемы, например заменяют «б» или «п» на «м». Другими словами, больные смешивают акустически разные, но близкие по артикуляции буквы. При этих поражениях коры страдает также и письмо. Больной может вместо слова «халат» написать «хадат» или «ханат». Считают, что такие симптомы являются результатом нарушения афферентной основы движения.
|
|
|
|
|
Дата добавления: 2014-10-15; Просмотров: 902; Нарушение авторских прав?; Мы поможем в написании вашей работы!