КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Экологические системы 2 страница
|
|
|
|
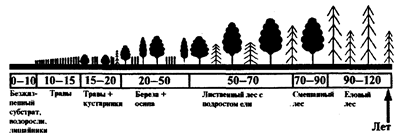
Однако прежде чем сформируется климаксное сообщество (экосистема), ему предшествует, как отмечалось выше, ряд промежуточных стадий или серий. Так, в лесной зоне (рис.5) на исходно безжизненном субстрате здесь сначала появляются организмы-пионеры, например, корковые водоросли, накипные лишайники и некоторые малотребовательные к плодородию субстрата семенные растения. За ними следует стадия растительности, представленная в основном травами, а затем кустарниками и деревьями-пионерами (чаще всего березой, осиной, ивой). Последние характеризуются быстрым ростом, но, отличаясь высоким светолюбием, быстро изреживаются (к 40-50-летнему возрасту). В результате этого под их пологом создаются условия для поселения теневыносливой ели, которая постепенно догоняет в росте стареющие лиственные виды деревьев и выходит в первый ярус. На данной стадии и образуется климаксное смешанное елово-лиственное сообщество или чисто еловый лес со свойственным им набором других видов растений и животных.
Рис. 5
Биогеоценотическая сукцессия на примере смен фитоценозов в лесной зоне
Наряду с теорией моноклимакса существует точка зрения, в соответствии с которой в одном и том же географическом районе может формироваться несколько завершающих (климаксных) экосистем (поликлимакс). Например, в лесной зоне наряду с еловыми и елово-лиственными лесами в качестве климаксных рассматривают также луговые экосистемы, сосновые леса. Однако сторонники моноклимакса считают, что луга в лесной зоне могут длительно существовать только в результате их использования (скашивания, выпаса). При прекращении таких воздействий на смену им неизбежно придут лесные сообщества. Что касается сосновых лесов, то длительное существование их связывается с тем, что они занимают обычно крайне бедные (например, песчаные, щебнистые, сильно заболоченные) местообитания, где ель (более сильный эдификатор) не может внедряться и существовать вследствие более значительной требовательности к почвенному плодородию. Но с течением времени по мере накопления в почве органических веществ и необходимых для жизни минеральных элементов и эти «сосновые местообитания», с точки зрения сторонников моноклимакса, будут заняты еловыми лесами, как обладающими более сильной эдификаторной способностью.
Причины сукцессий. Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделений или разложения и т. п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Например, в северных лесах внедрение ели под полог лиственных древесных сообществ связано прежде всего с тем, что она использует биологические свойства этих сообществ по слабому притенению почвы. Сами же почвенные условия остаются не только благоприятными для лиственных древостоев, но и постепенно улучшаются для них (идет накопление питательных веществ, уменьшается кислотность почв и т.п.). Следовательно, здесь нет оснований говорить о самоотравлении или других подобных причинах смен.
Не подтверждается безоговорочно и точка зрения о том, что появление ели под пологом лиственных древостоев связано с тем, что в молодом возрасте она требует затенения. Известно, например, что ель и в молодом возрасте прекрасно растет при полном освещении (значительно лучше, чем под пологом других древесных видов). Об этом, в частности, свидетельствуют многочисленные примеры создания культурных фитоценозов ели (посадкой молодых растений или посевом семян) на открытых площадях.
Наряду с природными факторами, причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). Преобладающие площади лесов представлены переходными (временными) экосистемами из лиственных древесных пород (береза, осина, реже ива, ольха и др.). Эти леса обычно называют производными или вторичными. Они, как отмечалось выше, являются промежуточными стадиями сукцессий.
К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т. п.
Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дигрессиями (лат. дигрессион - отклонение). Различают, например, пастбищные, рекреационные и другие дигрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложнение структуры, а стадиями катоценоза (греч. ката - вниз, против; кайнос - общий), которые нередко заканчиваются полным распадом экосистем.
Климаксные экосистемы обычно чувствительны к различным вмешательствам в их жизнь. К подобным воздействиям, кроме хвойных лесов, чувствительны и другие коренные сообщества, например, дубовые леса. Это одна из причин катастрофической гибели дубрав в современный период и замены их, как и хвойных лесов, менее ценными, но более устойчивыми временными экосистемами из березы, осины, кустарников или трав. Последнее особенно типично при разрушении степных и лесостепных дубрав.
Кроме песчаных пространств, первичные сукцессии могут начинаться на горных породах, извлеченных из недр, продуктах извержения вулканов (застывшая лава, отложения пепла) и т. п.
Вторичные н другие сукцессии. Вторичные сукцессии отличаются от первичных тем, что они начинаются обычно не с нулевых значений, а возникают на месте нарушенных или разрушенных экосистем. Например, после вырубок лесов, лесных пожаров, при зарастании площадей, находившихся под сельскохозяйственными угодьями. Основное отличие этих сукцессий заключается в том, что они протекают несравненно быстрее первичных, так как начинаются с промежуточных стадий (трав, кустарников или древесных растений-пионеров) и на фоне более богатых почв. Конечно, вторичная сукцессия возможна только в тех случаях, если человек не будет оказывать сильное и постоянное влияние на развивающиеся экосистемы. В последнем случае, как отмечалось выше, процесс пойдет по схеме дигрессий и завершится стадией катоценоза и опустынивания территорий.
Различают также автотрофные и гетеротрофные сукцессии. Рассмотренные выше примеры сукцессий относятся к автотрофным, поскольку все они протекают в экосистемах, где центральным звеном является растительный покров. С его развитием связаны смены гетеротрофных компонентов. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом, образующимися или фиксирующимися в организмах в процессе фотосинтеза либо хемосинтеза. Завершаются они, как отмечалось, климаксной стадией развития экосистем.
К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отсутствуют живые растения (продуценты), а участвуют лишь животные (гетеротрофы). Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества, в котором сменяются различные виды организмов-разрушителей. По мере разрушения органического вещества и высвобождения из него энергии сукцессионный ряд заканчивается, система распадается. Таким образом, эта сукцессия по природе своей деструктивна. Примерами гетеротрофных являются сукцессии, имеющие место, например, при разложении мертвого дерева или животного. Так, при разложении мертвого дерева можно выделить несколько стадий смен гетеротрофов. Первыми на мертвом, чаще ослабленном дереве поселяются насекомые-короеды. Далее их сменяют насекомые, питающиеся древесиной (ксилофаги). К ним относятся личинки усачей, златок и др. Одновременно идут смены грибного населения. Они имеют примерно следующую последовательность: грибы-пионеры (обычно окрашивают древесину в разные цвета), грибы-деструкторы, способствующие появлению мягкой гнили, и грибы-гумификаторы, превращающие часть гнилой древесины в гумус. На всех стадиях сукцессий присутствуют также бактерии. В конечном счете органическое вещество в основной массе разлагается до конечных продуктов: минеральных веществ и углекислого газа. Гетеротрофные сукцессии широко осуществляются при разложении детрита (в лесах он представлен лесной подстилкой). Они протекают также в экскрементах животных, в загрязненных водах, в частности, интенсивно идут при биологической очистке вод с использованием активного ила, насыщенного большим количеством организмов.
Общие закономерности сукцессионного процесса. Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса.
1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. По мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессии уменьшается вероятность вспышек численности отдельных видов.
4. Интенсифицируются процессы круговорота веществ, потока энергии и дыхания экосистем.
5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительны сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах.
6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обусловливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана.
7. В зрелой стадии климаксного сообщества (не старческой!) биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса. Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а во-вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов.
Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах (завершающие стадии сукцессий) по сравнению с лиственными (промежуточные стадии). Скорее, в последних больше потребителей зеленой продукции и вероятнее вспышки численности отдельных видов-фитофагов.
Нет также ни теоретических предпосылок,ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно (!) высоких значений.
Весь опыт лесоводства свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно-лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов.
Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная ее часть отчуждается гетеротрофами в результате образования устойчивых цепей выедания.
Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока, как правило, максимальна вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания консументов.
Стабильность и устойчивость экосистем
Термины «стабильность» и «устойчивость» в экологии обычно рассматриваются как синонимы, и под ними понимается способность экосистем сохранять свою структуру и функциональные свойства при воздействии внешних факторов.
Более целесообразно, однако, разграничивать эти термины, понимая под «стабильностью» данное выше определение, а под «устойчивостью» - способность экосистемы возвращаться в исходное (или близкое к нему) состояние после воздействия факторов, выводящих ее из равновесия. Кроме этого, для более полной характеристики реакции экосистем на внешние факторы целесообразно пользоваться в дополнение к названным еще двумя терминами: «упругость» и «пластичность».
Упругая система способна воспринимать значительные воздействия, не изменяя существенно своей структуры и свойств. Вместе с тем при определенных (запороговых) воздействиях такая система обычно разрушается или переходит в новое качество.
Пластичная система более чувствительна к воздействиям, но она под их влиянием как бы «прогибается» и затем относительно быстро возвращается в исходное или близкое к исходному состояние при прекращении или уменьшении силы воздействия.
Примером упругих экосистем являются климаксные (например, хвойные леса в лесной зоне, коренные тундровые сообщества, типчаково-ковыльные степи и т. п.). Пластичными экосистемами для лесной зоны являются лиственные леса как промежуточные стадии сукцессий. Они, например, выносят в несколько раз больше рекреационных (связанных с посещением населения) и других (пастьба скота, разного рода загрязнения) нагрузок, чем климаксные экосистемы, в которых эдификаторами выступают хвойные виды.
При рассмотрении стабильности и устойчивости как синонимов, обычно считается, что эти качества тем значительнее, чем разнообразнее экосистемы. Данное положение является настолько универсальным, что формулируется как закон: разнообразие - синоним устойчивости (автор Эшби). С этой точки зрения тундровые и пустынные экосистемы рассматриваются как малоустойчивые (нестабильные), а тропические леса, максимально богатые по видовому составу, - как самые устойчивые (стабильные).
Для экосистем с низкой устойчивостью характерны вспышки численности отдельных видов. Последнее связывается с тем, что в маловидовых экосистемах слабо проявляются силы, уравновешивающие численность различных видов (конкуренция, хищничество, паразитизм). Так, для тундровых экосистем типичны периодические резкие увеличения численности мелких грызунов - леммингов. В качестве результата низкой устойчивости этих экосистем рассматривается легкое разрушение их под влиянием внешних воздействий (перевыпаса, технических нагрузок и т. п.). Так, колеи, образующиеся после прохода тяжелой техники (тракторов, вездеходов), сохраняются десятилетиями.
С этих же позиций к неустойчивым и низкостабильным относят агросистемы, создаваемые человеком и представленные обычно одним преобладающим видом растений, интересующим человека. С этой же точки зрения как неустойчивые и нестабильные следует рассматривать сосновые леса на бедных песчаных или щебнистых почвах. Их древесный ярус представлен в таких условиях одним видом (сосной), беден в них и напочвенный (травяной, моховой) покров.
Однако если экосистемы, приведенные выше в качестве примеров, рассматривать с позиций названных выше различии устойчивости и стабильности, то они попадают в разные категории (табл. 4).
Устойчивость, стабильность и другие параметры экосистем зависят часто не столько от структуры самих сообществ (например, их разнообразия), сколько от биолого-экологических свойств видов-эдификаторов и доминантов, слагающих эти сообщества.
Так, высокая стабильность и значительная устойчивость, как видно из табл. 4, присущи сосновым лесам на бедных песчаных почвах, несмотря на малое видовое разнообразие этих экосистем. Это связано, во-первых, с тем, что сосна довольно пластична, и поэтому на изменение условий, например уплотнение почв, она реагирует снижением продуктивности и редко - распадом экосистемы. Однако и в последнем случае, в силу бедности субстрата питательными веществами и влагой, ее молодое поколение не встречает серьезной конкуренции со стороны других видов, и экосистема довольно быстро вновь восстанавливается в том же виде эдафического (почвенного) климакса.
Таблица 4
Характеристики устойчивости и стабильности отдельных экосистем
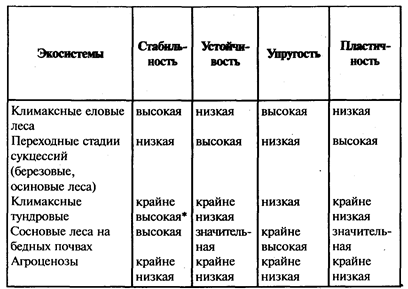
* по основным звеньям: фитоценозам и почвам
Иные параметры устойчивости и стабильности характерны для сосняков на богатых почвах, где они могут сменяться еловыми лесами, обладающими более сильными эдификаторными свойствами. Здесь, несмотря на значительное разнообразие (по видовому составу, ярусности, трофической структуре и т. п.), экосистемы сосновых лесов характеризуются низкой стабильностью и низкой устойчивостью. Сосна в данном случае выступает как промежуточная стадия сукцессионного ряда. Ей удается занимать и удерживать какое-то время такие местообитания только в силу каких-то необычных обстоятельств. Например, после пожаров, когда уничтожаются более сильные конкуренты (ель, лиственные древесные породы).
Классификация природных систем биосферы на ландшафтной основе
Классификация природных систем биосферы базируются на ландшафтном подходе, так как экосистемы - неотъемлемая часть природных географических ландшафтов, образующих географическую оболочку Земли.
Ландшафт – природный географический комплекс, в котором все основные компоненты, находятся в сложном взаимодействии, образуя однородную по условиям развития единую систему. По происхождению выделяют два типа ландшафтов – природный и антропогенный.
Наземные биомы (экосистемы)
В данном случае речь идет об экосистемах климатического климакса, т.е. развитые в них гомеостатические процессы наиболее совершенны в данных климатических условиях. Тундры характеризуются суровыми условиями (вечная мерзлота). Бореальные хвойные леса – тайга – распространены в северной части умеренной климатической. Листопадные леса умеренной зоны (широколиственные леса) расположены южнее тайги, не имеют сплошного распространения. Степи умеренной зоны – открытые пространства между лесами и пустынями в Евразии, в Северной Америке и Южной Америке, в Австралии и Новой Зеландии. Пустыни травянистые и кустарниковые располагаются в тропических и субтропических регионах в Азии, Африке, Австралии, Северной и южной Америках. Тропические степи и саванны – теплые области в Центральной и Восточной Африке, в Южной Америке и Австралии.
Пресноводные экосистемы
Составляя весьма малую часть от всех экосистем биосферы, пресноводные экосистемы для человека имеют непреходящее значение, вследствие того, что они практически единственный источник для бытовых и промышленных нужд. Пресные воды на поверхности континентов образуют реки, озера, болота, заполняют искусственные пруды и крупные водохранилища. Значит, пресные воды могут находиться в текучем, в относительно неподвижном стоячем и промежуточном состояниях.
Агроценозы
Основное свойство экосистем - способность естественного развития и прежде всего самовозобновления хотя бы в течение одного-двух поколений. С этой точки зрения нет основания рассматривать агроценозы как экосистемы или одну из стадий (начальную, промежуточную) сукцессионного ряда. Агроценозы сельскохозяйственных культур, особенно однолетних, существуют только при условии постоянного вмешательства человека. При прекращении такого вмешательства обычно начинается вторичная сукцессия с той стадии, которую называют сорняками. Но эта стадия уже не имеет прямого отношения к агроценозу.
Другими словами, такой агроценоз - это совершенно чуждое естественным условиям образование (сообщество), поэтому ему не присуще ни одно из свойств экосистемы
Несколько иные свойства характерны для агроценозов, создаваемых из долгоживущих лесных растений. Эти творения человека можно относить к экосистемам если не на протяжении всей жизни, то на определенных стадиях развития. Здесь наиболее типичны два варианта.
Первый из них относится к созданию леса искусственным путем в условиях, где сукцессии не являются четко выраженными. Например, сосняков на бедных песчаных почвах (см. выше). Здесь вмешательство человека по уходу за выращиваемыми растениями требуется только на начальных этапах жизни, когда сосенки еще настолько малы и слабы, что могут не выдержать конкуренции с травами. В дальнейшем (уже с 3-5 лет жизни) сосна начинает создавать сообщество и постепенно занимать позиции доминантного вида, формирующего свою среду. В последующем образуется сообщество со всеми критериями экосистемы. Правда, некоторые свойства такой экосистемы оказываются не вполне реализованными по сравнению с естественными сообществами. В частности, это проявляется в недостаточной жизнеспособности (устойчивости), обусловленной пониженным, по сравнению с естественными экосистемами, разнообразием. Последнее снижается в результате практически абсолютной выравненности растений по возрасту (возрастная структура как таковая отсутствует) и, в какой-то мере, выравненности почвенного фона в результате обработки почвы, предшествующей посадкам или посевам растений. Пониженная устойчивость проявляется через слабую дифференциацию деревьев по росту и, как следствие этого, ослабление, хотя и в разной степени, всех особей в период смыкания крон и острой внутривидовой конкуренции.
Кроме этого, равномерное размещение растений по площади и создание одновидовых древесных фитоценозов из хвойных видов способствует широко распространенному в настоящее время грибному заболеванию - корневой губке. Последняя имеет следствием очаговую, а порой и полную гибель растительных сообществ как систем.
Второй вариант связан с местопроизрастаниями (прежде всего почвами), характеризующимися значительным богатством питательными веществами и влагой. Здесь создание экосистем, минуя промежуточные стадии сукцессий, требует длительного вмешательства человека в их жизнь. Во всяком случае, до тех пор, пока вводимый вид (например, ель или сосна) не сформирует среду, препятствующую видам-конкурентам (осина, береза, ивы и др.) внедриться в сообщество и захватить главенствующие позиции. В большинстве же случаев победа оказывается на стороне естественных процессов развития экосистем (сукцессий). И виды, вводимые человеком, вытесняются конкурентами полностью или до такой степени, что они не способны сформировать полноценную экосистему с точки зрения целей, которые преследовал человек. Если удается сформировать желаемую экосистему (насаждение), минуя промежуточные стадии сукцессий (обязательно ценой больших затрат), то такие системы, как и в первом случае, оказываются недостаточно устойчивыми.
Исключить эти недостатки искусственных экосистем в значительной мере возможно посредством создания многовидовых сообществ, конечно, при постоянной помощи виду, в котором заинтересован человек. Иногда эти поправки удачно вносит природа через внедрение умеренного количества видов промежуточных стадий сукцессий (береза, осина и др.).
Таким образом, попытки создать сразу климаксные сообщества или их подобие, минуя промежуточные, часто обречены на неудачу по разным причинам. Это должен учитывать человек при решении конкретных хозяйственных проблем. Приведенные выше примеры свидетельствуют, насколько разнообразны взаимосвязи в экосистемах, их зависимость от биотических, абиотических и антропогенных факторов, а также насколько обязателен творческий (системный) подход в каждом конкретном случае, даже при том условии, что выявлены какие-то общие (часто основополагающие) закономерности существования экосистем. Возможности моделирования и создания человеком экосистем во многом зависят не только от биологических свойств видов (в приводимых примерах растительных), но и от условий местопроизрастания. Несомненна также относительность и необходимость конкретизации таких основополагающих экологических постулатов, как «разнообразие - синоним устойчивости», неизбежность резкого снижения продуктивности экосистем в климаксной стадии, неоднозначность терминов «устойчивость» и «стабильность» и др.
Человек и экосистемы
Человек в конкурентной борьбе за выживание в окружающей среде начал строить свои искусственные экосистемы 10 000 лет назад, создав сельское хозяйство - растениеводство и животноводство. На современном этапе он для удовлетворения все возрастающих потребностей вынужден изменять и даже разрушать природные экосистемы, может, и, не желая того. Энергия – это изначальная движущая сила природных и антропогенных экосистем. Их энергетические ресурсы могут быть неисчерпаемы и исчерпаемы.
Сельскохозяйственные экосистемы (агроэкосистемы)
Агроэкосистемы создаются человеком для получения высокого урожая – чистой продукции автотрофов. Основные их отличия от природных следующие:
§ резко снижено разнообразие видов культивируемых растений и животного населения биоценоза;
§ Растения и животные, культивируемые человеком, неконкурентоспособны в борьбе с дикими видами без поддержки человека;
§ Агроэкосистемы получают, кроме солнечной, дополнительную энергию, субсидируемую человеком;
§ Чистая продукция удаляется из экосистемы и не попадает в цепи питания биоценоза;
§ Экосистемы полей, садов, пастбищ, огородов – это упрощенные системы, поддерживаемые человеком на ранних стадиях сукцессии и неспособны к саморегуляции.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 796; Нарушение авторских прав?; Мы поможем в написании вашей работы!