КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
II. Ранние этапы эволюции предков человека
|
|
|
|
- Австралопитековые – двуногие человекообезьяны.
Первые находки. Впервые наименование австралопитек появилось в научной литературе в связи с ископаемыми находками Раймонда Дарта в 1924 году. Дарт обнаружил в доломитовых отложениях Юго-Восточного Трансвааля (ныне ЮАР), недалеко от местечка Таунг, череп детеныша гоминоида 3-5 лет («бэби из Таунга»). Кости черепа обладали в большей степени «обезьяньими» чертами с очень незначительными проявлениями «человечьих» признаков в строении челюстей. Внутренняя емкость черепа также более соответствовала средним показателям у большинства ископаемых и современных человекообразных обезьян – 380-450 см3.
Австралопитек африканский (Australopithecus africanus)- так назвал свою находку Р. Дарт, определив возраст австралопитека в 1.7 – 2.0 млн. лет. В дальнейшем в ряде мест Южной Африки (Стеркфонтейн, Макапансгат, Сварткранс, Кромдраай) помимо костей черепа были найдены остатки посткраниального скелета австралопитеков (тазовые кости, кости стопы), по которым удалось установить способность к двуногой локомоции.
Начиная с 1959 г. остатки австралопитеков стали находить и в Восточной Африке. Первые восточноафриканские австралопитеки были обнаружены супругами Мэри и Луисом Лики в древнейшем слое (№ I) Олдувайского ущелья в Танзании. Череп ископаемого гоминоида был очень массивным с характерным («гориллоидным») саггитальным гребнем на своде, мощными челюстями и крупными зубами (из-за чего был назван в шутку «Щелкунчик»). Лики присвоили своей находке наименование «Зинджантропа бойсова» (Zinjanthropm boisei), что означало «восточноафриканский человек», так как рядом с костями черепа были обнаружены грубо обработанные камни. С этого времени Восточная Африка и особенно Восточноафриканская рифтовая зона стали местом большого числа находок ранних гоминид – известны местонахождения Олдувай, Летоли, Кооби-Фора, Илерет, Канапои, Лотагам, Лукейно, Чесованджа, Чемерон, Хадар, Омо, Бодо, Арамис и др.
Богатейший ископаемый материал (несколько сотен индивидов), накопленный за 80-летнюю историю африканской палеоантропологии, порождает постоянные дискуссии относительно выделения отдельных родов, видов и положения австралопитеков в систематике гоминид.
Систематика австралопитековых. Иногда австралопитековых выделяют в отдельное семейство или относят к понгидам (Зубов, 1964). В данном случае они будут рассматриваться как настоящие гоминиды. Предлагается следующая систематика (Харитонов, 2003):
Семейство – Гоминиды (Hominidae)
Подсемейство Australopithecinae - Австралопитеки
Род Australopithecus – Австралопитек
Виды: Australopithecus afarensis – А. афарский
Australopithecus africanus – А. африканский
Paranthropus (Australopithecus) robustus – П. мощный
Paranthropus (Australopithecus) boisei – П. бойсов и др.
Подсемейство Homininae - Люди
Род Homo – Человек
Виды: H. habilis – Человек умелый
H. erectus – Человек выпрямленный
H. neanderthalensis - Человек неандертальский
H. sapiens sapiens – Человек разумный разумный
Среди антропологов существуют разные представления о количестве видов в пределах рода Homo. Помимо приведенных выше некоторыми специалистами выделяются H. rudolfensis (Человек рудольфский), H. ergaster (Человек деятельный), H. heidelbergensis (Человек гейдельбергский), H. rhodesiensis (Человек родезийский).
Положение австралопитековых в семействе гоминид можно считать достаточно обоснованным: во-первых, некоторые виды австралопитеков, вероятно, участвовали в происхождении более поздних предков человека; во-вторых, довольно трудно провести границу, отделяющую австралопитековых от первых «настоящих» Homo.
Разнообразие австралопитековых. Для определения физического типа австралопитеков можно выделить основные черты: двуногость (бипедия), малый мозг, крупные зубы с толстой эмалью (мегадонтия), небольшие клыки, отсутствие выраженного комплекса признаков в строении верхних конечностей, связанных с производством искусственных каменных орудий. В то же время в зависимости от древности и биологической специализации морфологические характеристики могут довольно существенно варьировать.
Самые последние находки определили хронологические рамки существования всех известных типов австралопитеков от 7 до 1 млн.лет.
В целом австралопитеков можно условно разделить на три основные группы, различные по морфологии и сравнительно последовательно сменявшие друг друга во времени (Дробышевский, 2002).
Таблица 4. Основные типы австралопитеков
(Дробышевский, 2002; с сокр.)
| Названия | Регион | время, млн.л. |
| Ранние австралопитеки наиболее ранние по времени, морфология крайне мозаична и включает как «понгидные», так и «человеческие» черты, некоторые черты строения уникальны; от человекообразных обезьян их надежно отличают малые размеры клыков и довольно явственные признаки прямохождения | ||
| Сахелантроп чадский Sahelanthropus tchadensis,2002 | Чад | 6-7 |
| Оррорин тугенский Orrorin tugenensis, 2001 | Кения | 5,65-6,2 |
| Австралопитек рамидас синонимы: Ардипитек рамидас Ardipithecus ramidus, 1995 | Эфиопия | 3,9-5,5 |
| Австралопитек анамский Australopithecus anamensis, 1995 | Кения | 3,9-4,2 |
| Грацильные австралопитеки(рис. 1) более поздняя группа, отличающаяся относительно небольшими размерами скелета и зубочелюстного аппарата; четкой грани между ранними и грацильными австралопитеками нет, основные отличия – возраст и меньшее количество примитивных признаков у более поздних видов; от массивных грацильные австралопитеки довольно четко отличаются меньшей общей массивностью и пропорциями зубных рядов: у грацильных обычно передние зубы относительно задних достаточно большие, коренные зубы не гипертрофированы в размерах. Предполагаемые параметры: длина тела - 105-150 см, масса тела - 30-50 кг, объем мозга – 380-450 см3 | ||
| Австралопитек афарский Australopithecus afarensis, 1978, синонимы: Австралопитек эфиопский, Парантроп африканский и др. | Кения, Эфиопия | 2,5-3,9-4,4? |
| Кениантроп Kenyanthropus platyops,2001 | Кения, Эфиопия | 3,2-3,5 |
| Австралопитек бахрелгазали Australopithecus bahrelghazali, 1996 | Чад | 3,0-3,5 |
| Австралопитек африканский Australopithecus africanus, синонимы: Плезиантроп трансваальский и др. | Южная и Восточная Африка | 2,4-3,5 |
| Австралопитек гархи Australopithecus garhi, 1999 | Эфиопия | 2,5 |
| Массивные австралопитеки(рис. 1) наиболее специализированная группа; характерны повышенная массивность черепа и, особенно, челюстей; значительно развит рельеф черепа; постоянный признак - заклыковая мегадонтия (резцы и клыки заметно редуцированы в размерах, премоляры и моляры резко увеличены и имеют толстую эмаль; Предполагаемые параметры: длина тела - 150-160 (160-180) см, масса тела - 50 (60-80) кг, объем мозга – 400-550 см3 | ||
| Парантроп эфиопский Paranthropus aethiopicus, 1968 | Кения, Эфиопия | 2,3-2,6 |
| Парантроп бойсов Paranthropus boisei, 1959 Australopithecus boisei, Австралопитек бойсов синонимы: Зинджантроп бойсов | Восточная Африка | 1,1-2,3-2,5? |
| Парантроп мощный, Paranthropus robustus, 1939 синонимы: Australopithecus robustus, Австралопитек мощный | Южная Африка | 0,9-2,5 |
Детальное изучение морфологии всех известных в настоящее время видов австралопитеков дает возможность разобраться в сложнейших проблемах становления прямохождения, усиленного развития головного мозга и появления культуры у последующих гоминид
Прямохождение, как древнейшая система гоминизации, начало формироваться, по-видимому, уже у преавстралопитековых форм и достаточно хорошо прослеживается у самых ранних австралопитеков около 7 млн. лет назад. В наибольшей степени прямохождение сказывается на строении тазового пояса:
- происходит расширение подвздошной кости кпереди, усиливается ее средняя часть;
- укрепляются крестцово-подвздошные и тазобедренные сочленения и происходит их сближение;
- развиваются элементы мускульно-связочного аппарата, фиксирующие разгибание ноги в тазобедренном и коленном суставах.
У австралопитеков форма таза и тазобедренный сустав в целом были подобны человеческим, они обладали постоянной двуногой походкой, что принципиально отличало их от всех известных ископаемых и современных обезьян.
Мозг австралопитеков по абсолютным размерам соответствовал вариациям его массы у современных человекообразных обезьян. Индивидуальные значения объема мозга колебались от 300 до 570 см3 (табл. 3). Нет четкого представления об изменениях в структуре моз- 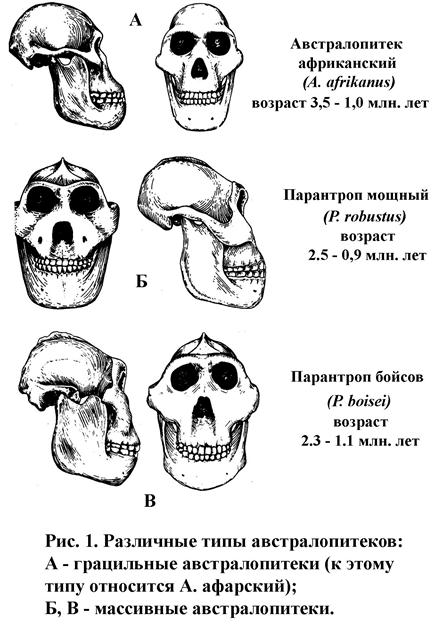 га (для этого используются эндокраны – внутренние слепки мозга). Существует мнение о понгидном типе строения мозга австралопитеков. В тоже время, отмечаются прогрессивные перестройки при сохранении малого объема самого мозга: увеличение теменной и височной ассоциативных зон.
га (для этого используются эндокраны – внутренние слепки мозга). Существует мнение о понгидном типе строения мозга австралопитеков. В тоже время, отмечаются прогрессивные перестройки при сохранении малого объема самого мозга: увеличение теменной и височной ассоциативных зон.
В строение черепа и зубной системы австралопитеков также имеется множество обезьяньих черт. Лицо было крупным с ярко выраженным прогнатизмом, подбородок отсутствовал, нос плоский и широкий, основание черепа слабо изогнуто, что свидетельствовало о примитивности голосового аппарата. Стадии развития постоянных зубов у австралопитеков были более сходны с аналогичными показателями у современных обезьян, чем у человека.
Важнейшая находка! В 1973 году в области Афарской депрессии на севере Эфиопии начала работу франко-американской экспедиция. В ноябре 1974 г. американский антрополог Д. Джохансон обнаружил миниатюрный скелет взрослой женской особи, названной впоследствии «Люси». Сохранность костных фрагментов составила около 40% всех костей скелета, что впервые позволило реконструировать черты внешнего облика древнейшего австралопитека и сделало находку уникальной (рис. 2) «Люси» было присвоено видовое название Australopithecus afarensis и определен ее возраст – более 3,5 млн. лет.
В морфологическом отношении «Люси» характеризуется целым комплексом примитивных особенностей. Двуногая ходьба не совсем соответствовала человеческой: меньшая длина шага, большой палец стопы был отведен сильнее, чем у человека, хотя и меньше, чем у шимпанзе. Кости малого таза «Люси» имели форму вытянутого поперечного овала на всех уровнях, что, возможно, свидетельствует об отсутствии поворота плода и более трудных, чем у обезьян, родах (результат прямохождения). В строении скелета еще прослеживаются следы адаптации к древесному передвижению: верхние конечности относительно длиннее нижних (хотя и не так, как у современных человекообразных), сохраняется изогнутость фаланг кисти и стопы. Полудревесный образ жизни «Люси» в пределах лесного биотопа подтверждается результатами пыльцевого анализа.
В строении черепа «Люси» также прослеживаются примитивные черты: малая емкость черепа -380-450 см3 (в среднем 413,5 см3); 
сагиттальные и затылочные гребни у предполагаемых мужских особей, сильное выступание лица, V-образная зубная дуга, крупные клыки, наличие диастем, и т.д. В соответствии с условной типологией (табл. 4) «Люси» может быть отнесена к грацильным австралопитекам (рис. 1).
Таким образом, «Люси» помогла установить, что по многим признакам афарский австралопитек (к этому виду относятся и некоторые последующие находки) соответствует представлению о «переходном звене» между понгидными предками и ранними представителями последующих гоминид.
Ранние австралопитеки (табл. 4), датируемые 7-6 млн. лет и представленные весьма фрагментарным костным материалом, могут рассматриваться как одни из возможных предков афарских австралопитеков.
Массивные австралопитеки (рис. 1), самые «молодые» представители австралопитековых, обитали в Восточной и Южной Африке в хронологическом интервале от 2,5 до 0,9 млн. лет. Вероятно, ведущей тенденцией развития этих гоминид было общее увеличение размеров тела, физической мощи, усиление жевательного аппарата при сохранении средних (для всех австралопитековых) показателей развития мозга. У массивных австралопитеков выработались специальные приспособления к эффективному и сильному жеванию: увеличенный саггитальный гребень на своде черепа, мощные челюсти с развитым костным рельефом (Парантроп бойсов – «Щелкунчик»), большая площадь жевательной поверхности моляров и премоляров и т.д. Такие морфологические особенности свидетельствуют о преобладании растительноядности. По-видимому, отсутствие потребности в мясной пищи не стимулировало развитие охоты, орудийной деятельности и мозга. Не совсем ясны причины отсутствия такой потребности: возможно, сильная конкуренция со стороны уже охотящихся австралопитеков грацильного типа и/или хищников «заставила» массивных австралопитеков специализироваться на употреблении растительной пищи. Вероятно, «пассивный» способ адаптации был одной из причин вымирания массивных австралопитеков около 1 млн. лет назад. Они представляли, видимо, тупиковую ветвь в эволюции ранних гоминид и разделили общую судьбу многих гигантских форм ископаемых животных, в том числе и некоторых приматов (гигантские лемуры Мадагаскара, гигантопитеки Юж. и Вост. Азии).
Среда обитания австралопитековых. Экологические условия, на фоне которых протекала эволюция австралопитековых в течение более 6 млн. лет, довольно существенно менялись. В Африке общее похолодание того времени сказалось на постепенном уменьшение влажности и изменении ландшафтов на более открытые и сухие. В тоже время, влажные и переменно-влажные леса все еще занимали обширные территории. Существенное уменьшение площади лесов и увеличение саванновых ландшафтов в Южной и Восточной Африке происходило с 3,6 млн. лет - время появления многих видов грацильных австралопитеков. Около 3 млн. лет площадь лесных биотопов опять увеличивается. Но, начиная с 2,5 млн. лет – время существования массивных австралопитеков - происходит значительное увеличение территорий саванн и редколесий и уменьшение лесов.
Данные палеоэкологии (анализ пыльцы, макрофауны, осадочных пород) позволяют реконструировать африканские ландшафты периода существования австралопитековых. Они чрезвычайно разнообразны, и в экологическом отношении можно говорить о «непрерывном ряду биотопов, на одном конце которого находится сезонный лес, а на другом – лесистая и открытая травяная саванна» (Фоули, 1990). Все же отмечается преобладание местонахождений австралопитековых в пределах саванновых редколесий и кустарниковых саванн. Самые сухие условия из всех известных в Африке характеризуют местонахождение массивного австралопитека в Пенинже (Танзания), где ландшафт представлял открытую травяную саванну.
«Остеодонтокератическая культура». В дословном переводе – культура кости – зуба – рога. Впервые была выделена Р. Дартом на основе находок многочисленных костных остатков австралопитеков рядом с большим количеством костей животных (антилоп, павианов). Некоторые кости были как будто обожжены, на черепах павианов – вмятины от ударов тяжелыми предметами. По мнению Дарта удары могли быть нанесены берцовыми костями антилоп (своеобразные палицы). Изучая форму расколотых костей из «кухонных куч», Дарт предположил, что австралопитеки использовали кости, рога и зубы в качестве орудий – «ножей», «кинжалов», намеренно их обрабатывая. Каменных орудий вместе с остатками австралопитековых пока не найдено. Отнесение природных орудий австралопитеков к категории «культура» условно. Теория Р. Дарта не получила всеобщего признания и, в значительной степени, была основана на интуиции исследователя (цит. по Зубову, 2002).
|
|
|
|
|
Дата добавления: 2014-11-16; Просмотров: 3616; Нарушение авторских прав?; Мы поможем в написании вашей работы!