КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Рефлекторный уровень организации движений 2 страница
|
|
|
|
Нейроэпителий утрикулюса лежит горизонтально, а саккулярный - парасагиттально. В результате этого утрикулярные рецепторы чувствительны к небольшим наклонам головы от ее нормального положения и к линейным ускорениям, которые возникают во время перемещения в горизонтальной плоскости. Саккулярные единицы в отличие от этого чувствительны к дорсовентральному ускорению, как это имеет место при прыжках и падениях.
В противоположность широким вариациям размеров тела у разных животных размеры полукружных каналов примерно одинаковы, также как размеры компаса мало зависят от размера корабля.
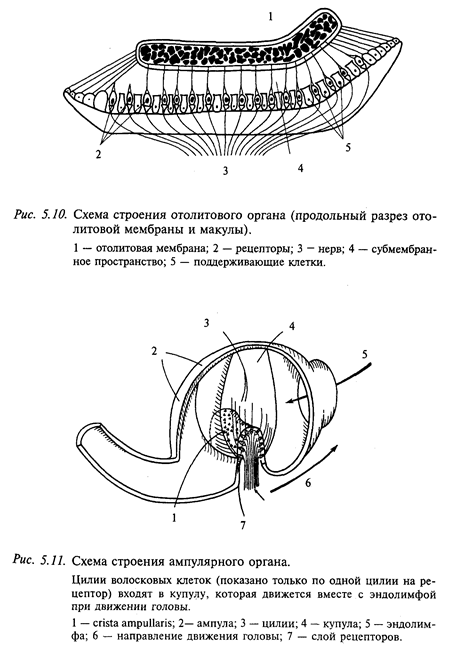
Волосковые клетки в нейроэпителии как отолитовых органов (саккулюсе и утрикулюсе), так и в ампулах полукружных каналов расположены упорядоченными рядами. Благодаря этому, например, отклонение купулы в сторону канала в горизонтальных каналах тормозит спонтанный разряд рецепторов, а в обратную сторону-увеличивает. В вертикальных каналах ориентация волосковых рецепторов противоположна: отклонение в сторону утрикулюса является тормозным, а в сторону канала - возбуждающим. В физиологических пределах усиление импульсации кристы при ускорении изменяется как линейная функция этого ускорения. Таким образом, каждый полукружный канал регистрирует степень ускорения в своей плоскости. В связи с тем что все три канала практически ортогональны (перпендикулярны относительно друг друга), они передают в ЦНС точную информацию об амплитуде и направлении углового ускорения головы в пространстве.
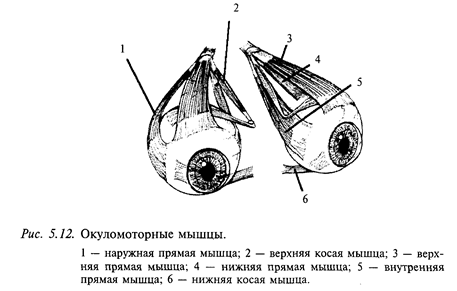
Краткие сведения об устройстве периферического звена глазодвигательной системы. Глазное яблоко управляется тремя парами мышц: двумя парами прямых и двумя косыми (медиальной и латеральной) (рис. 5.12). Каждая пара работает по принципу «тяни-толкай». Как уже указывалось, на каждой стороне черепа имеются три полукружных канала и отолитовые органы, представленные утрикулюсом и саккулюсом. Полукружные каналы одной стороны образуют прямоугольную систему координат. Каналы противоположных сторон, лежащие в одной плоскости, взаимодействуют между собой по принципу «тяни-толкай»: при вращении головы один канал пары максимально возбуждается, а другой максимально тормозится. Таким образом, все полукружные каналы образуют три пары: правый - левый горизонтальные, левый передний - правый задний и левый задний - правый передний. Каждая пара каналов связана с определенной парой глазодвигательных мышц трехнейронной цепочкой: горизонтальные каналы проецируются к горизонтальным прямым мышцам, левый передний - правый задний - к левой вертикальной прямой и правой косой, правый передний - левый задний каналы - к правой вертикальной и левой косой.
Установочные рефлексы мозгового ствола. Даже таламическое животное (т.е. животное, у которого удалены большие полушария) способно принимать и поддерживать нормальное положение тела. Это обеспечивается совокупным действием ряда рефлексов мозгового ствола. Примеры этих рефлексов приведены ниже.
1. Лабиринтный установочный рефлекс на голову состоит в том, что при любом положении тела голова животного принимает нормальное положение с параллельно расположенной ротовой щелью благодаря рефлексу с вестибулярного аппарата.
2. Тонические лабиринтные рефлексы на глаза. Компенсаторные движения глаз при поворотах головы и туловища можно наблюдать у всех позвоночных. Однако у животных с боковым положением глаз (например, у кролика) они выражены более отчетливо. Под действием этих рефлексов глаз максимально перемещается назад, когда голова животного занимает положение мордой вниз. Максимальный поворот глаза верхним полюсом роговицы вверх происходит при положении головы мордой вертикально вверх. Благодаря этим рефлексам каждому положению головы в пространстве соответствует определенное положение глазного яблока в орбите. При этом не имеет значения, с какой стороны достигалось положение головы. Всегда устанавливается одно и то же закономерное положение глаз. У животных с фронтальным расположением глаз (например, у хищных, приматов) при наклоне головы вперед глаза отклоняются вверх. Тонические реакции глаз рефлекторно возникают в результате раздражения отолитового аппарата.
При повороте головы кролика вокруг сагиттальной плоскости также возникают тонические лабиринтные рефлексы на глаза. При этом отклонения обоих глаз при разных положениях головы в пространстве происходят в противоположных направлениях. Если вращать голову от нормального положения (ротовая щель расположена горизонтально), то глаз, находящийся внизу, отклонится в орбите вверх. Отклонение достигает максимума при положении головы на боку. Контралатеральный глаз при этом отклонится вниз. При вращении через положение на затылке реакция глаз быстро изменяется на обратную.
Таким образом, существуют два независимых компонента тонического лабиринтного рефлекса на глаза, которые образуют все возможные комбинации при каждом положении головы в пространстве. Быстрые движения глаз, поскольку они вызываются с полукружных каналов, не участвуют в этих статических рефлексах. Активирование полукружных каналов (вращением головы) вызывает вестибулоокулярный рефлекс.
3. Вестибулоокулярный рефлекс (ВОР) - один из древнейших; он сопровождает позвоночных животных на протяжении всей их эволюции. Рефлекс проявляется в том, что при вращении головы глаза в орбитах противовращаются. Внешне это проявляется в виде нистагма колебательных движений глазных яблок в орбитах. Различают медленную компоненту нистагма, при которой глаза вращаются в сторону вращения головы примерно с той же скоростью, и быструю фазу нистагма, при которой глаза в орбитах после достижения своего крайнего положения скачком (саккадой) возвращаются в исходное положение. Этот рефлекс можно наблюдать как в полной темноте, так и на свету. Биологическая роль этого рефлекса - предотвращать сползание сетчаточного изображения при вращении головы. Входным сигналом для ВОР является импульсация, передаваемая по вестибулярным афферентам от полукружных каналов. Вестибулярные афференты направляются в мозговой ствол (верхнее вестибулярное ядро).
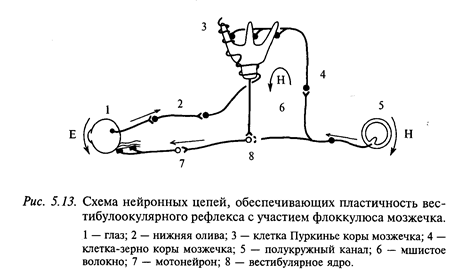
Этот афферентный сигнал после обработки выходит на эффекторное звено экстраокулярные мышцы. Главная проблема для такой системы состоит в калибровке: как отрегулировать скорость противовращения глазных яблок, чтобы оно эффективно компенсировало вращение головы. На рис. 5.13 показана принципиальная схема ВОР. В процессе калибровки скорости противовращения глазных яблок в орбитах относительно направления вращения головы участвует мозжечок. Это символически показано одной клеткой коры мозжечка (клеткой Пуркинье), а именно - самой задней частью мозжечка (флоккулонодулярная), которую часто называют вестибулярным мозжечком (см. приложение 6).
Гипотеза об участии флоккулюса мозжечка в пластичности ВОР была высказана на основе тесных связей вестибулярного мозжечка (flocculus и nodulus) как с вестибулярной, так и со зрительной системой. В пользу этого предположения говорят также результаты по удалению вестибулярного мозжечка. Оказалось, после удаления вестибулярного мозжечка (флоккулюса и парафлоккулюса), а также нижней половины VIII дольки и нижних 3-4 долек парамедианной доли усиление ВОР устанавливалось на уровне 1,063 (диапазон 0,95-1,173) и исчезала способность ВОР к пластическим изменениям коэффициента усиления.
.
Предполагают, что сигналом для изменения усиления ВОР является смещение сетчаточного изображения. Далее этот сигнал по дополнительной зрительной системе поступает в претектальную область (ядро оптического тракта), затем к нижней оливе и через лазающие волокна из нижней оливы к клеткам Пуркинье коры вестибулярного мозжечка (флоккулюсу). Сигнал скорости вращения головы передается от верхнего вестибулярного ядра по мшистым волокнам к клеткам-зернам и далее через параллельные волокна поступает также на клетки Пуркинье. Таким образом, клетки Пуркинье коры вестибулярного мозжечка выполняют оль компараторов (сравнителей), благодаря «работе» которых может модифицироваться коэффициент усиления ВОР.
У человека ВОР участвует в реакции установки взора. Эта реакция возникает при появлении сбоку в поле зрения неожиданного зрительного стимула. В первый момент человек скачкообразно переводит глаза в сторону стимула и «захватывает» зрением этот стимул. Слово «захватывает» означает, что стимул помещается в область сетчатки с наилучшим разрешением (зрительная ямка, fovea). С небольшим отставанием (это связано с инерцией) в эту же сторону начинает перемещаться голова. Чтобы изображение не сползло с сетчатки (другими словами, чтобы человек не потерял изображение), включается ВОР, который в данном случае запускается благодаря вращению головы и, следовательно, возбуждению рецепторов полукружных каналов. При этом скорость противовращения глаз в орбитах равна скорости вращения головы. Благодаря этому взор (алгебраическая сумма скоростей вращения глаз и головы) остается неподвижным в пространстве.
ВОР обеспечивает движение глаза в противофазе с движениями головы в диапазоне 0,01-6,5 Гц. На более низкой частоте в стабилизации сетчаточного изображения участвуют также оптомоторный рефлекс и рефлекс прослеживания. Под «стабилизацией» будем понимать то, что положение глаза относительно зрительной цели не меняется, несмотря на вращение головы. Это достигаете механизмом ВОР: глаза противовращаются в орбитах со скоростью, равной скорости вращения головы.
Для успешной стабилизации сетчаточного изображения при вращениях головы необходимо регулировать коэффициент усиления ВОР. Нетрудно себе представить ситуации, в которых необходимо изменять усиление ВОР. Например, при росте организма меняются размеры головы и глазных яблок, в силу разных обстоятельств (травмы, процессы старения и пр.) меняется упругость глазодвигательных мышц и многое другое. Коэффициент усиления ВОР регулируется в коре задней части мозжечка.
4. Оптомоторный рефлекс проявляется в том, что при движении в поле зрения человека структурированного зрительного поля происходит непроизвольное движение глазных яблок в сторону движения стимулов. Когда движение глазных яблок в орбитах исчерпывает свой диапазон (по 45 угл. градуса от среднего положения), глаза скачкообразно возвращаются в исходное состояние. Совместная работа трех систем (вестибулоокулярной, оптомоторной и прослеживания) обеспечивает латентный период реакции глазного яблока, не превышающий 80 мс. Другими словами, человек способен следить за объектами, движущимися со скоростью около 300 угл. град. в секунду.
5. Шейные рефлексы на глаза. Если у животного (например, у кролика) удалить лабиринты, а затем поворачивать его голову, то можно получить все те перемещения глаз, которые описаны выше. Но в данном случае движения глаз осуществляются исключительно за счет шейных рефлексов. Положение глаз, вызванное этими рефлексами (при изменении положения головы или туловища), в конечном итоге приводит к тому, что изображение на сетчатках обоих глаз остается стабильным. В норме это достигается только совместным действием лабиринтных и шейных рефлексов. Шейноокулярный рефлекс довольно плохо выражен у интактных взрослых млекопитающих и практически отсутствует в норме у человека.
6. Шейный рефлекс на конечности. Если поднимать или опускать голову животного в атлантоокципитальном суставе, то вызываемые при этом рефлексы на передние и задние конечности действуют противоположным образом: при опускании головы активируются флексоры передних конечностей и экстензоры задних, при поднимании головы активируются экстензоры передних конечностей и флексоры задних (см. рис. 5.8). Сравнивая тонические рефлексы, можно видеть, что шейные и лабиринтные рефлексы на передние конечности действуют как синергисты, а на задние - как антагонисты.
7. Рефлекс перераспределения тонуса конечностей при поворотах головы. Если голову децеребрированной кошки (удалены большие полушария), лежащей на животе, поворачивать, то повышается тонус экстензорной мускулатуры в «челюстных» конечностях (конечности той половины туловища, к которым поворачивается нос), уменьшается и увеличивается - в «черепных» (конечности, к которым в этом случае поворачивается череп).
Такое перераспределение тонуса для четвероногих животных оправдано соответствующим перенесением центра тяжести, и для того чтобы выдержать увеличенный вес в соответствующих конечностях, увеличивается их разгибательный (экстензорный) тонус. Можно получать все описанные шейные тонические рефлексы, если фиксировать позвоночный столб и перемещать только переднюю часть шеи.
У здорового взрослого человека шейные и лабиринтные рефлексы не проявляются, но они отчетливо выступают в патологических состояниях (гидроцефалия, опухоли мозга, тяжелые мозговые травмы и пр.). Например, в этих условиях обнаруживаются шейные тонические рефлексы. Если поворачивать голову влево, чтобы нос приближался к левому плечу, то левые конечности становятся «челюстными», а правые - «черепными», поэтому повышается разгибательный тонус в обеих конечностях левой половины тела и уменьшается-справа.
8. Оптический установочный рефлекс играет особенно важную роль в правильной установке головы у высших млекопитающих. Например, обезьяна с разрушенным лабиринтом может правильно устанавливать голову, но если надеть ей колпак, закрывающий глаза, т.е. лишить зрения, она уже не сможет правильно устанавливать голову в пространстве.
Таким образом, в организме животных взаимодействует ряд установочных рефлексов, благодаря которым обеспечивается нормальное положение головы и тела. Описанные установочные рефлексы осуществляются нервными центрами на уровне не выше среднего мозга.
9. Лифтная реакция представляет собой рефлекс на прямолинейное ускорение. Рефлекс состоит в том, что при поднятии площадки, на которой находится животное, тонус экстензоров снижается, и конечности подгибаются; при опускании площадки развивается противоположная реакция: экстензоры активируются и конечности разгибаются. Эта реакция легко вызывается и у человека при подъеме или опускании в скоростном лифте.
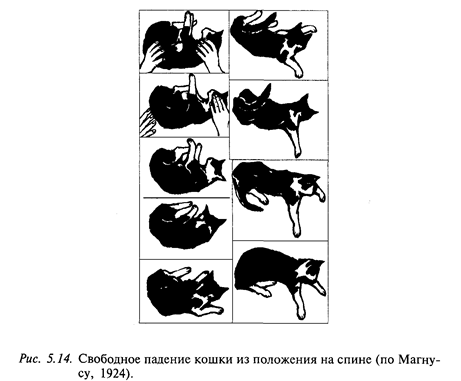
Описанные выше установочные рефлексы легко наблюдать на интактной кошке, которая свободно падает из положения на спине (рис. 5.14). Прежде всего, активируются лабиринтные рефлексы на голову, благодаря которым она поворачивается по направлению к нормальному положению. К этой реакции присоединяется шейный установочный рефлекс, вследствие чего за головой следует тело, сначала грудной отдел и только затем таз. Происходит своего рода штопорообразное движение животного в пространстве, начинающееся с головы. При этом передние конечности разгибаются. Дальнейшее прямолинейное смещение головы в пространстве при свободном падении вызывает разгибание задних конечностей. Последнее можно объяснить лабиринтным рефлексом на падение (линейное ускорение). Благодаря совокупному действию этих рефлексов голова и тело кошки принимают нормальное положение, конечности тонически разгибаются и готовы принять вес тела, когда животное достигнет земли.
Таким образом, положение тела и головы в пространстве и по отношению друг к другу принадлежит к наиболее застрахованным и автоматически выполняемым реакциям ЦНС. Благодаря взаимной деятельности большого количества разнородных рефлексов осуществляется единая конечная цель - правильная установка головы и тела в пространстве. Статокинетические и статические реакции взаимно дополняют друг друга: благодаря кинетическим реакциям выполняется движение, приводящее отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами.
Глазодвигательная система млекопитающих. В эволюции наземных позвоночных животных возникла необходимость сохранять неподвижность изображения на сетчатке не только в состоянии покоя или медленных движений, но также во время локомоции. Эволюция глазодвигательной системы у этих животных происходила в тесном взаимодействии с эволюцией зрительной системы. С точки зрения эволюции глазодвигательной системы всех позвоночных животных можно условно разделить на две группы: «фовеальные» животные, сетчатка которых имеет fovea - и они обладают достаточно острым зрением, и «афовеальные» животные - с недостаточно острым зрением. У «фовеальных» животных (к ним относится и человек) глаза расположены во фронтальной плоскости, у них есть бинокулярное зрение. Зрительная ямка, или fovea,-это небольшая область в сетчатке с повышенной плотностью фоторецепторов, причем только в этой области находятся колбочки - фоторецепторы дневного (и цветового) зрения. Глазодвигательная система млекопитающих, которая сформировалась как необходимое дополнение зрительной системы, позволяет рассматривать зрительный мир. С точки зрения физиологии непрерывно решается задача - движением глаз поместить на зрительную ямку наиболее значимые части изображения. Что будет, если отключить fovea? Это происходит с каждым из нас при снижении уровня освещенности, например, вечером. Известно, что мы в этих условиях не видим мелких предметов, не можем читать. Для такого зрения движения глаз не обязательны. Действительно, у животных с невыраженной fovea, например собаки, коровы, быстрые движения глаз отсутствуют.
Глазодвигательная система млекопитающих выполняет следующие задачи: 1) сохраняет неподвижным изображение внешнего мира на сетчатке во время движения животного относительно этого мира; 2) выделяет во внешнем мире некоторые объекты, помещает их в зоне сетчатки с высоким разрешением (например, fovea у человека) и прослеживает их движениями глаз и головы; 3) осуществляет скачкообразные (саккадические) перемещения взора для сканирования (рассматривания) внешнего мира. «Афовеальные» животные, например кошка, собака, кролик и др., не заботятся о положении изображения на сетчатке, и вестибулярная и оптокинетическая системы только предотвращают скольжение изображения по сетчатке. У человека в связи с изложенными выше требованиями, кроме вестибулоокулярной и оптокинетической систем, имеется также система прослеживания, вергенции и саккад. У него развились также рефлекторные механизмы содружественного движения глаз и головы.
Саккадная система в полной мере развита только у обезьян и человека. Именно у них есть бинокулярное зрение, и есть «фовеальное» зрение.
Саккады - это быстрые содружественные отклонения глаз в быструю фазу вестибулярного и оптомоторного нистагмов, начальная фаза реакции прослеживания, когда скачком глаза «захватывается» движущаяся зрительная цель при зрительном обследовании внешнего мира. Структуры мозга, обеспечивающие быстрые, скачкообразные движения глаз - саккады, находятся в ретикулярной формации мозгового ствола. Однако высший центр произвольного управления движениями глаз локализован в коре лобной доли мозга и называется фронтальным глазодвигательным полем (8-е и 10-е поля).
Активность нейронов мозгового ствола. Схема, представленная на рис. 5.15, показывает связи и вероятные события, приводящие к саккаде глаза. Важными элементами этой схемы являются пачечные нейроны и нейроны-паузы. Во время фиксации взора клетки-паузы разряжаются тонически с высокой частотой; так как тормозные связи от них идут к тормозным и возбудительным пачечным нейронам, это должно предотвращать формирование пачки импульсов. Предполагают, что возбудительные пачечные нейроны проецируются к тоническим нейронам и происходит интегрирование их активности, в результате чего образуется сигнал, необходимый для поддержания глаза в эксцентричном положении (так называемый нейронный интегратор). В действительности нейронный интегратор, по-видимому, имеет большую протяженность. Установлено, что разрушение в области мозгового ствола и мозжечка приводит к параличу взора, т. е. неспособности поддерживать эксцентричное положение взора.
По логике схемы стволового генератора запускающий сигнал формируется клетками-паузами. Установлено, что они получают афферентацию из средних и глубоких слоев верхнего двухолмия. Электрофизиологическими методами также было показано, что клетки-паузы могут возбуждаться моносинаптически на электрическую стимуляцию нижних слоев верхнего двухолмия. Таким образом, нейронная схема генератора саккады «работает» по принципу «спускового крючка» - клетка-пауза постоянно тормозит пачечные нейроны. Как только из верхнего двухолмия затормаживается разряд клетки-паузы, в этот интервал растормаживается пачечный нейрон и этот сигнал подается на соответствующие мотонейроны, что и приводит к саккаде.
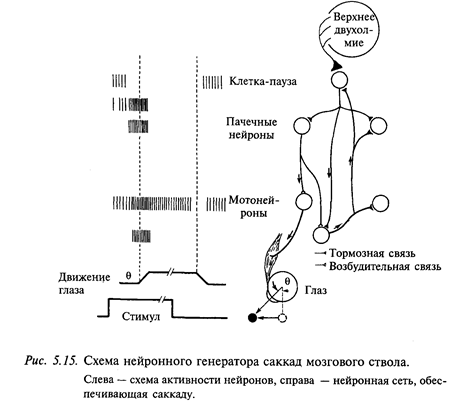
На основании электрофизиологических данных было высказано предположение, что пачечные нейроны со спонтанным разрядом являются промежуточным звеном между нижними слоями переднего двухолмия и парамедианной ретикулярной формацией моста. По мнению исследователей, эти нейроны могут получать конвергентные входы из ряда источников, в том числе из фронтального глазодвигательного поля. Известно, что дефицит саккад возникает только при одновременном разрушении переднего двухолмия и фронтального глазодвигательного поля.
|
|
|
|
|
Дата добавления: 2014-11-08; Просмотров: 533; Нарушение авторских прав?; Мы поможем в написании вашей работы!