КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Виды торможения в центральной нервной системе
|
|
|
|
Понятие о процессах возбуждения и торможения в центральной нервной системе.
Возбуждение и торможение представляют собой два противоположных по сути, но в то же время взаимосвязанных и взаимодополняющих физиологических процесса, обеспечивающих функционирование нервной системы. Возбуждение проявляется в деполяризации мембраны нейрона, что может сопровождаться генерацией нервного импульса в области его аксонного холмика, дальнейшим распространением нервного импульса по аксону и передачей через возбуждающие синапсы на другие нейроны или структуры рабочего органа. Благодаря тому, что каждая чувствительная или вставочная нервная клетка вступает в синаптический контакт с большим количеством других нейронов, возбуждение способно к распространению (иррадиации) в центральной нервной системе. Именно возбуждение обеспечивает передачу информации в центральной нервной системе и возможность дальнейшего проведения этой информации к исполнительным органам, включая их в деятельность или изменяя исходный (базальный) уровень их активности. Процесс возбуждения в нервной системе связан с деятельностью возбуждающих синапсов (в центральной нервной системе. высших позвоночных в основном возбуждающих синапсов химического типа и значительно реже – электрических синапсов). В возбуждающих синапсах химического типа выделяемый вследствие деполяризации пресинаптической мембраны медиатор вызывает деполяризацию постсинаптической мембраны (генерацию возбуждающего постсинаптического потенциала) путем увеличения ее проницаемости для ионов натрия или кальция. Как правило, на мембране нейронов имеется большое количество синапсов (сотни - тысячи), образуемых другими нейронами; среди этих синапсов могут быть контакты как возбуждающего, так и тормозного типа. В случае одновременного прихода нервного импульса к пресинаптическим окончаниям различных синапсов одного и того же нейрона (т.е. по различным афферентным входам нейрона) в разных участках его мембраны (которые являются постсинаптическими мембранами для этих синапсов) возникают постсинаптические потенциалы (возбуждающие и тормозные в зависимости от типа синапсов), которые могут суммироваться в пространстве. При этом возбуждающие постсинаптические потенциалы соседних участков мембраны нейрона будут усиливать друг друга, а тормозные постсинаптические потенциалы, возникающие на постсинаптической мембране тормозных синапсов, напротив, будут ослаблять суммарный возбуждающий постсинаптический потенциал нейрона. В случае ритмического следования нервных импульсов к пресинаптическому полюсу одного и того же синапса какого-то нейрона возникающие последовательно с определенным интервалом времени на постсинаптической мембране этого синапса постсинаптические потенциалы (как возбуждающие так и тормозные) будут суммироваться во времени. Это означает, что каждый следующий нервный импульс, приходящий к пресинаптическому полюсу, будет усиливать постсинаптический потенциал, вызванный предыдущим нервным импульсом, если частота прихода импульсов к пресинаптическому окончанию такова, что постсинаптическая мембрана за межимпульсный интервал не успевает полностью реполяризоваться.
Торможение в центральной нервной системе, как правило, связано с деятельностью тормозных синапсов, вызывается возбуждением, не способно к распространению, всегда локализуется в определенных участках центральной нервной системы, и его назначение состоит в подавлении или ослаблении другого возбуждения. Несмотря на то, что торможение является процессом, противоположным по сути возбуждению, оно неотъемлемо физиологически связано с ним, поскольку способствует локализации возбуждения в определенных участках центральной нервной системы, включению в конкретные рефлекторные реакции не всех, а только лишь строго определенных нейронов. Выключение или ослабление центрального торможения приводит к невозможности нормального функционирования нервной системы. Так, например, введение в организм лягушки стрихнина (блокирует функцию тормозных синапсов) сопровождается нарушением центрального торможения и, как следствие этого, любой действующий на организм раздражитель приводит к генерализованной ответной реакции животного, затрагивающей многие органы и различные скелетные мышцы, в норме в данной реакции не участвующие. В частности, при ударе по столу вблизи животного, которому ввели стрихнин, вместо обычного прыжка от места нанесения раздражения, будут наблюдаться судороги многих скелетных мышц, изменение сердечной деятельности, интенсивности дыхания и некоторые другие не характерные в норме реакции.
Впервые явление центрального торможения было открыто И.М. Сеченовым в опытах на лягушках в 1862 г. Сущность опыта состояла в следующем: лягушке с удаленными большими полушариями головного мозга накладывали на таламические бугры кристаллы поваренной соли (раздражая их таким образом) и при этом наблюдали ослабление некоторых спинальных рефлексов (рефлексов, замыкающихся на уровне спинного мозга). В частности, у животного резко ослабевал рефлекс одергивания нижней конечности при помещении ее в раствор соляной кислоты. В дальнейшем Ф. Гольц показал, что торможение в центральной нервной системе можно вызвать не только путем непосредственного раздражения определенных ее отделов (подобно тому, как в опыте И.М. Сеченова раздражением таламуса), но и другими способами. Так, например, подавить или ослабить сгибательный рефлекс одной из лапок лягушки при помещении этой лапки в раствор кислоты можно, не только раздражая таламическую область, но и нанося сильное механическое раздражение (сдавление) на другую конечность. Таким образом, исследованиями Ф. Гольца, был доказан тот факт, что торможение в центральной нервной системе не обязательно связано с деятельностью особых тормозных центров, а может развиваться в любом отделе центральной нервной системы.
В зависимости от механизмов, обеспечивающих центральное торможение, выделяют следующие его виды:
Ø постсинаптическое торможение; обусловлено деятельностью тормозных синапсов в центральной нервной системе. В данном случае поступление нервного импульса к пресинаптической мембране тормозного синапса, образуемого каким-то одним нейроном (тормозным нейроном) на определенных структурах другого нейрона, приводит к гиперполяризации постсинаптической мембраны (возникновению тормозного постсинаптического потенциала), что ослабляет суммарную деполяризацию соседних участков нейрона, на которых могут быть расположены возбуждающие синапсы, также активируемые в данный момент. Причем эффективность такого постсинаптического торможения во многом зависит от локализации тормозного синапса на нейроне. Так, наиболее эффективное тормозное влияние в целом на всю нервную клетку оказывают тормозные синапсы, расположенные вблизи области аксонного холмика, где, как правило, возникает нервный импульс в ответ на суммарную деполяризацию мембраны тела и дендритов нейрона, развивающуюся в результате прихода информации к нейрону по различным его возбуждающим синапсам. Возникновение же тормозного постсинаптического потенциала вблизи аксонного холмика будет ослаблять суммарный возбуждающий постсинаптический потенциал мембраны тела нейрона и может сделать невозможной генерацию нервного импульса в области аксонного холмика. Таким образом, в случае постсинаптического торможения распространение нервного импульса к пресинаптическому окончанию тормозного нейрона, образующего тормозный синапс на другом нейроне, приводит к возникновению гиперполяризации (тормозного постсинаптического потенциала) на участке мембраны иннервируемого нейрона, т.е. к ее непосредственному торможению.
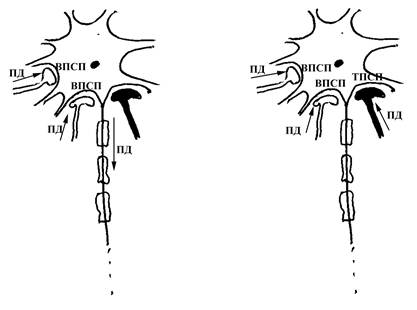
А Б
Рис. 8. Схема, объясняющая механизм постсинаптического торможения. Обратите внимание на то, что на мембране тела нейрона расположено несколько синапсов (на данном рисунке три синапса, причем два из них (незатушеванные) являются возбуждающими, а один тормозным (затушеванный синапс)). На рисунке А показан такой вариант, когда возбуждение (в виде нервного импульса или потенциала действия, ПД) приходит только к пресинаптическим полюсам возбуждающих синапсов и соответственно на постсинаптической мембране этих синапсов (являющейся частью мембраны тела нейрона) возникают возбуждающие постсинаптические потенциалы (ВПСП, проявляющиеся в локальной деполяризации постсинаптической мембраны). Такие ВПСП, одновременно возникающие на постсинаптической мембране соседних синапсов, суммируются в пространстве (взаимоусиливают друг друга) и возникающий суммарный постсинаптический потенциал является достаточным для инициации ПД в области аксонного холмика данного нейрона. На рисунке Б показан такой вариант, когда возбуждение в виде нервного импульса одновременно приходит к преснаптическим полюсам всех трех синапсов (двух возбуждающих и одного тормозного); при этом на постсинаптической мембране тормозного синапса (являющейся частью мембраны тела рассматриваемого нейрона) возникает тормозный постсинаптический потенциал (который может проявляться в виде гиперполяризации постсинаптической мембраны), и такой потенциал ослабляет суммарный возбуждающий постсинаптический потенциал, возникающий в результате активации двух других возбуждающих синапсов, до такой степени, что генерация ПД в области аксонного холмика нейрона становится невозможной. Постсинаптическое торможение будет тем более эффективным для нейрона, чем ближе располагаются тормозные синапсы по отношению к аксонному его холмику.
Ø пресинаптическое торможение; обеспечивается всегда аксо-аксональными химическими синапсами тормозного типа. В частности, аксон какого-то нейрона (как правило, тормозного нейрона) или коллатераль аксона (его боковое ответвление) образует тормозный синапс на пресинаптическом окончании аксона другого нейрона. Медиатор, выделяемый в таком тормозном синапсе, увеличивает проницаемость постсинаптической мембраны (которая в свою очередь является частью пресинаптического окончания другого нейрона) для ионов Cl-. При этом хлор сразу после открытия соответствующих хлорных каналов в постсинаптической мембране тормозного синапса движется внутрь постсинаптического полюса по концентрационному градиенту, гиперполяризуя мембрану. Но затем, когда концентрация хлора внутри постсинаптического полюса становится выше таковой в синаптической щели, ионы хлора начинают, напротив, выходить из постсинаптического полюса, приводя к стойкой и длительной деполяризации постсинаптической мембраны тормозного синапса. Отмеченное сопровождается инактивацией потенциалзависимых натриевых каналов в пресинаптической терминали аксона иннервируемого нейрона, снижением ее возбудимости и нарушением проведения нервного импульса к пресинаптической мембране этой терминали. Таким образом, в случае пресинаптического торможения посредством аксо-аксональных тормозных синапсов блокируется проведение возбуждения (нервного импульса) еще на пути к какому-то нейрону (на уровне пресинаптических терминалей возбуждающих нейронов, вступающих в контакт с данным нейроном).
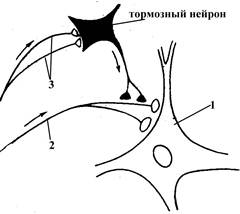 |
Рис. 9. Схема осуществления пресинаптического торможения в нервной системе (по Бабскому Е.Б, 1972). Тормозный нейрон (изображен черным) образует тормозные синапсы не на самом теле нейрона, на который оказывается его тормозное влияние (на рисунке обозначен цифрой 1), а на пресинаптических терминалях аксона (обозначены цифрой 2) другого нейрона, оказывающего возбуждающее действие на нейрон 1. Таким образом, в случае пресинаптического торможения тормозные нейроны оказывают не непосредственное тормозное воздействие на другой нейрон, а опосредованное, блокируя проведение к нему возбуждения по аксонам иннервирующих его возбуждающих нейронов. Сам же тормозный нейрон активируется в результате прихода к нему возбуждения по нервным волокнам, обозначенным на рисунке цифрой 3.
Ø торможение по типу пессимума Введенского (пессимальное торможение) может возникать в обычных возбуждающих синапсах при слишком частом следовании к ним нервных импульсов. В таких условиях медиатор, выделяющийся при каждой предыдущей деполяризации пресинаптической мембраны, не успевает расщепиться соответствующим ферментом, и концентрация медиатора в области постсинаптической мембраны с каждым последующим нервным импульсом, приходящим к пресинаптическому полюсу, нарастает. Стойкая же и длительная деполяризация постсинаптической мембраны приводит к натриевой инактивации в околосинаптической мембране иннервируемого нейрона и снижению ее возбудимости
Ø торможение вслед за возбуждением развивается в некоторых нейронах по той причине, что нервный импульс, возникающий в области аксонного их холмика, сопровождается очень длительной следовой гиперполяризацией. При возникновении длительной следовой гиперполяризации возбудимость мембраны аксонного холмика снижается, и она оказывается временно неспособной генерировать новые потенциалы действия в ответ на суммарную деполяризацию, возникающую на мембране тела и дендритов данного нейрона при поступлении к нему информации через различные возбуждающие афферентные входы.
Понятие о рефлексе и рефлекторной дуге. Классификации рефлексов
В основе деятельности нервной системы лежат рефлексы. Рефлекс – это ответная реакция организма на действие раздражителя, осуществляемая при обязательном участии нервной системы. Наиболее простой пример рефлекса: прикосновение руки к горячему предмету инициирует почти моментальное ее одергивание. В данном случае в качестве раздражителя выступает высокая температура ощупываемого объекта, в качестве рецепторов, воспринимающих действие раздражителя, – терморецепторы кожи, в качестве рабочего органа – мышца-сгибатель верхней конечности, а в качестве ответной реакции – сокращение этой мышцы и удаление руки от места нанесения раздражения.
Путь рефлекса (его материальная основа, т.е. совокупность нервных элементов и определенных структур рабочего органа, взаимодействие между которыми обеспечивает осуществление рефлекса) представляет собой его рефлекторную дугу. Любая рефлекторная дуга состоит из следующих компонентов:
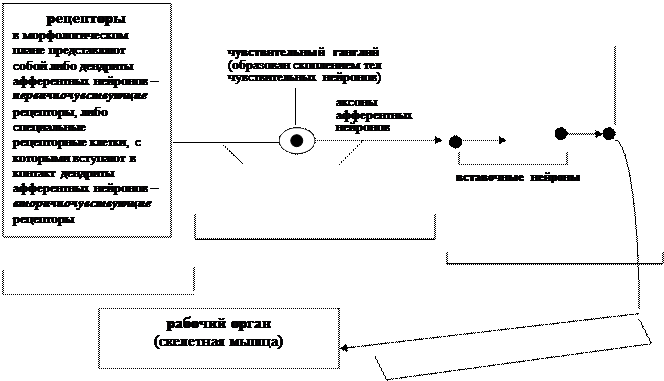
Ø рецепторов – специализированных воспринимающих структур, расположенных, как правило, на периферии (в органах чувств, коже, внутренних органах, скелетных мышцах, сосудах и т.д.) и морфологически представленных дендритами чувствительных нейронов (первичночувствующие рецепторы) или специализированными рецепторными клетками ненервной природы, с которыми вступают в контакт дендриты чувствительных нейронов (вторичночувствующие рецепторы);
Ø афферентного звена, представленного телами и аксонами афферентных нейронов, обеспечивающих проведение информации от рецепторов в центральную нервную систему;
Ø нервного центра, представленного вставочными нейронами, заложенными в определенных участках центральной нервной системы, а, возможно, и на разных ее уровнях, и перерабатывающих афферентную информацию;
Ø эфферентного звена, представленного эфферентными нейронами, передающими ответную программу действий ("эфферентную" информацию) к исполнительным органам;
Ø исполнительных органов, включение в деятельность или изменение деятельности которых обеспечивает ответную реакцию организма на действие раздражителя.
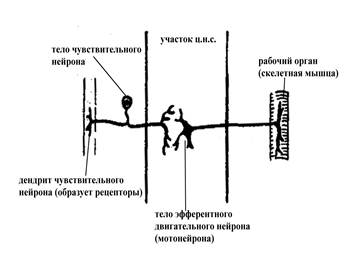 Самые простые рефлекторные дуги не имеют вставочного звена, и информация от чувствительного нейрона непосредственно передается на эфферентный, иннервирующий определенный исполнительный орган (такие рефлекторные дуги называют двухнейронными моносинаптическими, чтобы подчеркнуть наличие всего лишь одного синаптического переключения на пути нервного импульса от чувствительного нейрона на эфферентный). Рефлекторные дуги большинства рефлексов характеризуются наличием вставочного звена, которое может быть различной степени сложности и разветвленности (многонейронные полисинаптические рефлекторные дуги).
Самые простые рефлекторные дуги не имеют вставочного звена, и информация от чувствительного нейрона непосредственно передается на эфферентный, иннервирующий определенный исполнительный орган (такие рефлекторные дуги называют двухнейронными моносинаптическими, чтобы подчеркнуть наличие всего лишь одного синаптического переключения на пути нервного импульса от чувствительного нейрона на эфферентный). Рефлекторные дуги большинства рефлексов характеризуются наличием вставочного звена, которое может быть различной степени сложности и разветвленности (многонейронные полисинаптические рефлекторные дуги).
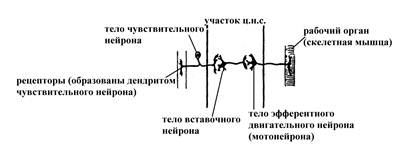
А Б
Рис. 10. Упрощенные схемы моносинаптической двухнейронной (А) и полисинаптической многонейронной (Б) рефлекторных дуг (по Бабскому Е.Б, 1972).
 | |||||
 | |||||
 |
|
|
|
|
|
|
|
|
|
|
|
|
 | |||||
 | |||||
| |||||
|
|
|
|
|
|
|
|
|
|
|
|
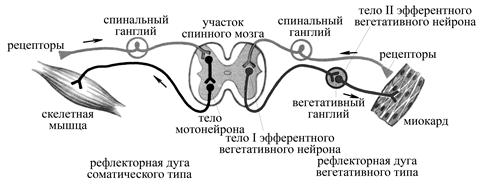 Рис. 12. Схема рефлекторной дуги вегетативного типа.
Рис. 12. Схема рефлекторной дуги вегетативного типа.
Рис. 13. Схема рефлекторных дуг соматического и вегетативного рефлексов, замыкающихся на уровне спинного мозга (по Коробкову А.В., Чесноковой С.А., 1987).
Рефлексы можно классифицировать на основании:
Ø их биологического значения для организма (пищевые, оборонительные, локомоторные, позно-тонические, половые, ориентировочные и т.д.);
Ø локализации рецепторов, раздражение которых вызывает данный рефлекс (экстероцептивные (рецепторы этих рефлексов заложены в коже, органах чувств и воспринимают изменения во внешней среде), висцероцептивные (рецепторы заложены во внутренних органах, сосудах и воспринимают изменения во внутренней среде организма), проприоцептивные (рецепторы заложены в скелетных мышцах))
Ø того, на уровне каких отделов центральной нервной системы замыкаются рефлекторные дуги рефлексов (спинальные (на уровне спинного мозга), бульбарные (на уровне продолговатого мозга), мезэнцефальные (на уровне среднего мозга), диэнцефальные (на уровне промежуточного мозга), кортикальные (на уровне коры больших полушарий))
Ø того, являются ли рефлексы врожденными (безусловные) или приобретенными (условные) реакциями организма
Ø от того, какие структуры организма выступают в качестве рабочего органа рефлекса: вегетативные рефлексы (в качестве рабочего органа выступают внутренние органы, сосуды, железы или гладкомышечные волокна кожи) и соматические рефлексы (в качестве рабочего органа выступают скелетные мышцы). Рефлекторные дуги вегетативных и соматических рефлексов отличаются строением своего эфферентного звена. Так, в рефлекторной дуге соматического типа на пути нервного импульса из центральной нервной системы к рабочему органу находится один эфферентный нейрон, тело которого заложено в самой центральной нервной системе, а аксон покидает ее и вступает в контакт со скелетными мышечными волокнами. В рефлекторной дуге вегетативного типа на пути нервного импульса из центральной нервной системы к рабочему органу (внутренние органы, железы и гладкомышечные волокна кожи, гладкая мускулатура сосудов) имеется два эфферентных нейрона: тело первого заложено в самой центральной нервной системе, его аксон (преганглионарное нервное волокно) выходит из центральной нервной системы и переключается на тело второго эфферентного вегетативного нейрона, заложенного в вегетативном узле, аксон же второго эфферентного вегетативного нейрона (постганглионарное нервное волокно) вступает в контакт со структурами иннервируемого органа.
Сложные рефлекторные акты, в которых завершение одного рефлекса обуславливает возникновение другого и им сменяется, носят название цепных рефлексов. Многие инстинкты (пищевой, половой, родительский и др.) осуществляются по типу цепных рефлексов. Одной из разновидностей цепных рефлексов являются ритмические рефлексы, представляющие собой чередование в определенной последовательности одних и тех же простых рефлекторных актов, которые ритмически повторяются. Примером ритмических рефлексов являются рефлекс шагания, почесывания и другие.
|
|
|
|
|
Дата добавления: 2014-11-16; Просмотров: 5403; Нарушение авторских прав?; Мы поможем в написании вашей работы!