КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Возрастные изменения ЭЭГ и ВП
|
|
|
|
О
GD
Со
младспчестло зрелость млаяекчеепш зрелость В
младенчество зрелость младенчество зрелость С

младепчестцо зрелость младенчество зрелость Рис. 17.2. А, В, С — варианты моделей психогенетики развития.
Генетическая вариативность (обозначена кругами) и генетическая ковариация между младенчеством и зрелостью (обозначены перекрытием кругов) [по; 355].
оценить вклад генетических факторов в изменчивость возрастных преобразований, а также установить, насколько связаны между собой генетические и средовые компоненты межиндивидуальной вариативности признака в разные периоды. Иначе говоря, для того чтобы судить о преемственности или стабильности генетических и средовых влияний, необходимо лонгитюдное исследование близнецов или сиб-сов, которое позволит установить степень связи между генетическими компонентами (межвозрастная генетическая корреляция) и средовы-ми компонентами (межвозрастная средовая корреляция) фенотипи-ческой дисперсии признака.
Таким образом, полная схема исследования в психогенетике развития с необходимостью включает эмпирическое исследование и наследуемости, и генетических корреляций. Сама по себе высокая наследуемость признака, полученная в разных возрастах, ни в коей мере не свидетельствует о стабильности генетических влияний.
По представлениям Р. Пломина, эти две переменные — наследуемость и степень генетической общности, определяемая величиной генетической корреляции, — относительно независимы и могут образовывать разные варианты сочетаний (рис, 17.2), Модель А, например, предполагает, что наследуемость признака с возрастом может оставаться без изменений (левый столбик) или возрастать (правый), но независимо от этого генетической преемственности при этом не обнаруживается, т.е. генетические эффекты в раннем возрасте и зрелости абсолютно не связаны между собой. В моделях В и С, напротив, допускаются варианты частичной (В) или (С) полной генетически опосредованной преемственности в формировании признака. Последний вариант (с точки зрения Р. Пломина, наиболее вероятный) получил название амплификационной модели. Амплификационная мо-
дель реализации генетических влияний в онтогенезе предполагает, что с возрастом, по мере созревания ЦНС и формирования индивидуально устойчивых способов переработки информации, возрастает наследуемость признака, при этом сохраняется высокая межвозрастная генетическая корреляция, т.е. генетические эффекты, действовавшие в младенчестве, высоко коррелируют с генетическими эффектами в зрелости.
Более детализованную модель предлагает Л. Ивс с соавторами [245]. В зависимости от времени начала экспрессии генов они выделяют две альтернативы в генотипической детерминации развития. Первая предполагает, что все гены находятся в действенном состоянии с момента рождения и развитие есть модификация фенотипа средовыми влияниями. В этом случае наследуемость признака в онтогенезе будет уменьшаться, приближаясь к некой асимптотической величине, которая есть функция исходной наследуемости и «условной памяти», ответственной за фиксацию средового опыта. Вторая исходит из того, что гены постоянно синтезируют продукты, требуемые для информационной обработки. В таком случае наследуемость будет возрастать от небольшой величины в момент рождения до высокой асимптотической величины, которая является функцией исходной или первоначальной наследуемости и постоянства экспрессии генов во время развития.
При анализе данных лонгитюдного исследования близнецов модель Ивса предполагает, что фенотип каждого индивида во времени О, 1, 2...t выступает как функция аддитивных генетических эффектов и уникального средового опыта. В каждый момент времени имеются новые генетические эффекты и новые специфические средовые влияния. Если принять, что все средовые эффекты являются случайно-специфическими, а генетические эффекты действуют через интервенцию генного продукта, который может сохраняться в течение времени, то генетические эффекты на фенотип в данный момент времени есть результат генов, экспрессирующихся вновь вместе с эффектами генов, которые экспрессировались на всех предшествующих стадиях развития в той степени, в какой они сохраняются во времени. По мере развития признака генетические эффекты будут накапливаться, приводя к увеличению генотипической и фенотипической вариативности.
Таким образом, модель Ивса в простейшем случае (когда генетические эффекты постоянны во времени, средовые — случайно специфичны, а пути распространения влияний постоянны), как и ампли-фикационная модель Пломина, предсказывает увеличение генотипи-ческого компонента фенотипической вариативности в онтогенезе. Однако, анализируя межвозрастные связи между генетическими составляющими вариативности, Ивс прогнозирует уменьшение генетического компонента ковариации между двумя фенотипами одного и того же индивида в разные моменты времени u и t, причем генетический компонент ковариации будет уменьшаться как экспоненциальная функция интервала времени u — t.
В моделях Пломина и Ивса в центре анализа находится структура фенотипической дисперсии и рассматривается онтогенетическая динамика в формировании индивидуальных особенностей, при этом генетический анализ динамики средних значений изучаемых характеристик, как таковых, остается за пределами внимания авторов.
Однако существует модель, в которой органически сочетается генетический анализ лонгитюдных средних и ковариационной структуры. Она базируется на использовании авторегрессионной симплексной модели [202, 258], Не вдаваясь в детали математического аппарата, отметим, что эта модель позволяет экспериментально выяснить, одни и те же или разные генетические и средовые факторы объясняют фенотипическую вариативность и фено-типические средние. Фактически данная модель впервые на экспериментальном уровне ставит проблему взаимосвязи генотип-средовой детерминации нормативных характеристик и их индивидуальных различий.
5. ВОЗРАСТНАЯ ДИНАМИКА ГЕНЕТИЧЕСКИХ
И СРЕДОВЫХ ДЕТЕРМИНАНТ В ИЗМЕНЧИВОСТИ КОГНИТИВНЫХ ХАРАКТЕРИСТИК
Представления о том, что в онтогенезе меняется соотношение генетических и средовых влияний, определяющих индивидуально-психологические особенности, родились в контексте самой психогенетики, тем не менее они хорошо согласуются с представлениями возрастной психологии, касающимися изменения механизмов реализации психических функций в онтогенезе.
Выдающийся отечественный психолог А,Р. Лурия писал: «Мы имеем все основания думать, что природа каждой психической функции (иначе говоря, ее отношение к генотипу) так же меняется в процессе психического развития человека, как и ее структура, и что поэтому ошибочными являются попытки раз и навсегда решить вопрос «о степени наследственной обусловленности» той или иной психической «функции», не принимая в расчет тех изменений, которые она претерпевает в своем развитии» [99].
На основе теоретических представлений, существующих в отечественной возрастной психологии, делались попытки определить направление этих изменений. Так, А. Р. Лурия, исходя из фактов качественной перестройки всей психической деятельности человека и замены элементарных форм деятельности сложноорганизованными функциональными системами, которые происходят в процессе психического развития, предположил, что по мере изменения структуры высших психических функций, возрастания степени их опосредования зависимость той или иной деятельности от генотипа будет закономерно уменьшаться. Действительно, исследования некоторых особенностей памяти и внимания выявили именно такой характер изменений. Было установлено, что от дошкольного к подростковому возрасту сохраняется преимущественно генотипическая обусловлен-
ность образной зрительной памяти и устойчивости внимания. В то же время наблюдается фактическая смена детерминации, т.е. переход от генотипической к средовой обусловленности у опосредованных форм памяти и у более сложных форм внимания, таких, как его распределение [2, 97]. Наряду с этим исследования генотип-средовых отношений в показателях интеллекта в ходе развития дают другую картину.
Возрастная стабильность и изменчивость генетических и средовых влияний, лежащих в основе межиндивидуальных различий по интеллекту, в последнее время являются предметом многих исследований [25, 56, 106, 355].
В большинстве исследований делается вывод о том, что в младенчестве наследственная обусловленность показателей интеллекта относительно низка, а влияние систематической семейной среды сравнительно велико. Начиная с шести лет и далее, а также у подростков и взрослых оценка наследуемости показателей интеллекта возрастает до 50-70%, влияние же общей семейной среды существенно снижается (подробнее об этом см. гл. VI).
Эти выводы представляют обобщение результатов целого ряда исследований, выполненных на близнецах и приемных детях. Рассмотрим сначала результаты некоторых близнецовых исследований.
Наиболее известным из них является Луисвиллское близнецовое исследование, посвященное изучению природы межиндивидуальной изменчивости показателей интеллекта. Оно было начато в 1957 г. Ф. Фолкнером и к середине 80-х годов охватывало около 500 пар близнецов, чье развитие было прослежено от рождения до 15-летнего возраста. Близнецы, участвовавшие в этом исследовании, тестировались по интеллекту, начиная с первого года жизни до 15 лет (каждые три месяца на протяжении первого года жизни, дважды в год — до 3 лет, ежегодно до 9 лет и последний раз — в 15 лет). При этом использовалась шкала психического развития Бейли в младенческом периоде, шкалы Векслера — WPPSI в возрасте 4,5 и 6 лет и WISC в более старшем возрасте.
Анализ полученных оценок IQ в парах МЗ и ДЗ близнецов отчетливо демонстрирует увеличение показателя наследуемости с возрастом. Показатели наследуемости у детей в возрасте 1, 2, 3 лет составляют 10, 17, 18% соответственно. С 3 лет внутрипарное сходство МЗ близнецов сохраняется на очень высоком уровне, коэффициенты корреляции не ниже 0,83. У ДЗ близнецов внутрипарное сходство по показателю интеллекта уменьшается с 0,79 в 3 года до 0,54 в 15 лет. Показатели наследуемости у детей в 4 года составляют 26% и далее увеличиваются до 55%.
Интересно, что значимые различия в сходстве МЗ и ДЗ близнецов начали обнаруживаться до того, как была установлена их зиготность. Тип близнецовых пар, т.е. отнесение их к МЗ или ДЗ близнецам, был определен только в 3 года. По мнению исследователей, получен-
ные данные показывают, что процессы развития инициируются и в значительной степени управляются генотипом. Это предположение было подтверждено при изучении внутрипарного сходства индивидуальных траекторий, или профилей, развития МЗ и ДЗ близнецов,
Профиль индивидуального развития характеризует не только направление развития психологических характеристик, но и индивидуальные особенности движения в этом направлении, которые могут включать периоды ускорения и замедления, спада и подъема. В Луис-виллском близнецовом исследовании по результатам многолетнего прослеживания изменений в уровне интеллекта МЗ и ДЗ близнецов оказалось возможным провести внутрипарное сравнение профилей индивидуального развития по показателям интеллекта [452, 453].
Внутрипарное сравнение профилей показало, что в парах МЗ близнецов наблюдается значительно большее совпадение значений интеллекта по каждому году, т.е. наблюдается больше сходства по ходу развития в целом. В парах ДЗ близнецов совпадения были выражены значительно меньше (рис. 17,3). Мерой количественной оценки внутрипарного сходства профилей показателей IQ служат коэффициенты корреляции, которые составляют 0,87 для МЗ близнецов и 0,65 для ДЗ в возрастном диапазоне от 3 до б лет и 0,81 и 0,66 соответственно в диапазоне от 6 до 8 лет. Коэффициенты наследуемости равны соответственно 0,44 для первого возрастного отрезка и 0,30 — для второго. Таким образом, индивидуальные особенности профилей развития по показателям интеллекта испытывают на себе существенное влияние генотипа, однако вполне возможно, что степень этого влияния на разных отрезках онтогенеза также будет варьировать.
Как уже отмечалось, основным достоинством лонгитюдного исследования близнецов является то, что только оно может ответить на вопрос: обусловлено ли увеличение наследуемости проявлением новых дополнительных генетических факторов, начинающих функционировать по мере взросления ребенка, или происходит усиление уже действующих генетических факторов?
Л. Ивс с коллегами [245] подошел к анализу генотип-средовых соотношений в развитии интеллекта именно с этих позиций, используя для анализа материалы лонгитюдного Луисвиллского близнецового исследования. Анализ полученных у близнецов в разные годы оценок IQ выявил изначально небольшое, но устойчивое и возрастающее влияние одних и тех же генетических факторов. Было установлено и существенное влияние систематической семейной среды, причем эффекты систематической среды также сохраняли преемственность, хотя по мере взросления к стабильным присоединялись и новые. Несистематические средовые влияния были менее устойчивы по сравнению с генетическими и систематическими средовыми эффектами. В целом эти данные свидетельствуют в пользу амплификационной модели наследуемости показателей IQ.
Рис. 17.3. Профили развития МЗ и ДЗ близнецов [453]. а — МЗ близнецы; б — ДЗ близнецы.
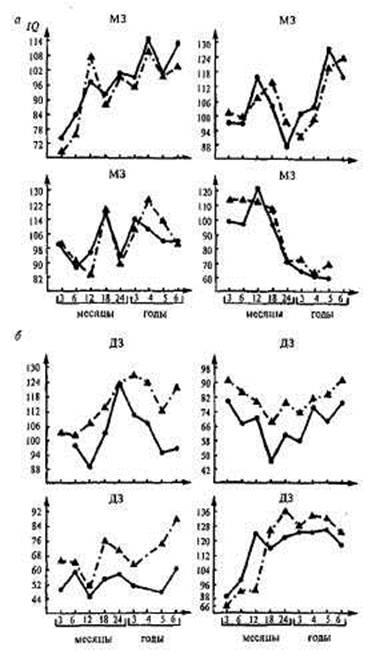
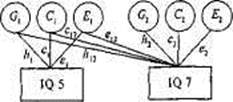
Рис. 17.4. Диаграмма путей двумерной генетической модели для одного признака (IQ) в 5 и 7 лет [25].
□ — измеряемые переменные; О—латентные, не наблюдаемые впрямую факторы. Gb и G2 — генетические влияния на IQ u 5 и 7 лет, Q и С2 — систематическая (общая) среда, E1 и Е2 — индивидуальные средовые факторы. Коэффициенты путей hb сь в\ и h2, с2, е2 представляют влияние латентных факторов на IQ во временные периоды 1 и 2 соответственно; коэффициенты путей h12, с12 и еи представляют относительное влияние G1, Q и E1, на IQ в 7 лет. У МЗ близнецов внутри-парная корреляция генотипов равна 1,0, у ДЗ близнецов — и среднем 0,5. Как у МЗ, так и у ДЗ внутрипарная корреляция систематической среды составляет 1,0; случайная (несистематическая) среда не скоррелирована.
Сходные проблемы решались и в ряде других подобных исследований. Так, голландские исследователи Д. Бумсма и К. Ван Баал [25] приводят результаты лонгитюдного генетического исследования IQ у близнецов 5-7 лет. Они ставили задачу проанализировать изменения соотношения средовых и генетических факторов, влияющих на IQ, и выделить компоненты фенотипической стабильности в указанном возрастном диапазоне. Для оценки интеллекта близнецов использовался один и тот же тест RAKIT (обновленный амстердамский тест детского интеллекта). Выборка испытуемых включала: в 5 лет 209 пар близнецов, в 7 лет повторно 192 пары. Генетико-статистический анализ проводился с привлечением генетической модели, которая в качестве источников вариативности рассматривала генетические эффекты, систематическую среду, несистематическую среду (рис. 17.4).
Сопоставление оценок наследуемости и их доверительных интервалов в 5 и 7 лет показало, что относительное влияние генетических факторов действительно различается в двух возрастах. Несмотря на то что в 5 лет верхняя граница наследственной обусловленности — 42 %, а в 7 лет ее нижняя граница — 50%, непересекающийся доверительный интервал свидетельствует о значительно более высокой наследуемости в 7 лет. Соответственно относительное влияние систематической среды ниже в 7 лет, чем в 5 лет. Корреляция между оценками интеллекта в 5 и 7 лет составила 0,65. В генетической модели ковариа-ция показателей IQ между 5 и 7 годами разлагалась на генетическую и средовую составляющие, при этом большая часть ее — 64% объясняется стабильностью генетических факторов, а меньшая — 36% — стабильностью систематической среды.
Таким образом, исследование Д. Бумсмы и К. Ван Баал показало, что, во-первых, наследуемость показателей IQ в возрастном интервале с 5 до 7 лет возрастает и, во-вторых, в основе межиндивидуальной изменчивости показателей IQ в обоих возрастах лежат преимущественно одни и те же генетические факторы. Следует указать, что данное близнецовое исследование в некотором роде уникально, так как наряду с оценкой когнитивных функций в нем проводился анализ межиндивидуальной изменчивости большого числа психофизиологических показателей, характеризующих особенности созревания ЦНС (описание этих данных см, в гл. XVIII). Перспективы подобного параллельного изучения представляются очень интересными, поскольку можно будет, фактически впервые, проанализировать природу межуровневых и межвозрастных связей в структуре развивающейся индивидуальности.
Изменения генотип-средовых соотношений в показателях интеллекта приблизительно в том же возрастном диапазоне изучались в близнецовом лонгитюдном исследовании, проводившемся М.С. Егоровой и ее коллегами [56]. Ставилась задача проследить динамику генотип-средовых соотношений в показателях интеллекта при переходе от дошкольного возраста к школьному. В исследовании приняли участие около 100 пар близнецов, Диагностика интеллекта проводилась по тесту Векслера (WISC), адаптированному А. Панасюком. Показатели наследуемости для общего интеллекта (ОИ) составили 28, 34 и 52% в 6, 7 и 10 лет соответственно; для вербального интеллекта (ВИ) — 22, 16 и 26%, для невербального (НИ) — 16, 84 и 70%. Эти данные интересны тем, что, с одной стороны, подтверждают тенденцию к возрастанию наследуемости общего интеллекта с возрастом, а с другой — свидетельствуют о возможности иных вариантов возрастных изменений наследуемости отдельных сторон интеллекта.
В этом же исследовании анализировались межвозрастные генетические корреляции, позволявшие оценить генетический вклад в фе-нотипическую стабильность показателей интеллекта (табл. 17.2).
Генетические корреляции между показателями вербального, невербального и общего интеллекта в 6 и 7 лет, а также генетические корреляции между этими показателями в 6 и 10 лет за небольшим исключением достаточно высоки. В то же время генетические корреляции между всеми показателями интеллекта в 7 и 10 лет намного ниже. Таким образом, генетические факторы в изменчивости интеллекта в 6 лет достаточно тесно связаны с генетическими факторами, обусловливающими индивидуальные различия в 7 и 10 лет. Авторы обращают внимание на то, что «выпадение» из общей картины корреляций между показателями в 7 и 10 лет может быть обусловлено резкими изменениями средовых условий, связанных с началом обучения в школе.
В целом описанные близнецовые исследования убедительно свидетельствуют о весьма существенных возрастных изменениях в карти-
Таблица 17.2
Генетические корреляции между суммарными показателями интеллекта [56]
| Возраст | Показатели интеллекта | 6 лет | 7 лет | 10 лет | ||||||
| ВИ | НИ | ОИ | ВИ | НИ | ОИ | ВИ | НИ | ОИ | ||
| 6 лет | ВИ НИ ОИ | 95 84 | 40 56 | 77 62 | 61 63 | 24 42 | 42 43 | |||
| 7 лет | ВИ НИ ОИ | 95 92 | 25 -12 11 | 12 46 | 22 08 19 | |||||
| 10 лет | ВИ НИ ОИ |
Примечание. Здесь и в табл. 17.3, 18.1 нули и запятые опущены.
не генотип-средовых соотношений в изменчивости показателей IQ. Более того, очевидны и дальнейшие перспективы близнецовых лон-гитюдных исследований, связанные с более дробным дифференцированным анализом генетических и средовых влияний в показателях когнитивных характеристик разного уровня и содержания, оценкой их стабильности и преемственности в онтогенезе.
Наряду с близнецовыми проводятся лонгитюдные исследования приемных детей. Среди них наиболее известным является Колорадское исследование приемных детей, которое было начато по инициативе Р. Пломина и Дж. Дефриза в 1975 г. [361]. В исследовании принимали участие 246 семей с детьми первого года жизни. По мере ежегодного тестирования число семей уменьшалось, и к девятилетнему возрасту детей оно составило 173. Предполагается продолжать исследование до того времени, когда детям исполнится 16 лет.
У детей, начиная с первого года жизни, ежегодно диагносциро-вали показатели физического и умственного развития по шкалам Н. Бейли. На основании наблюдений и оценок родителей делались выводы об особенностях темперамента ребенка, (О возрастной динамике генотип-средовых соотношений в индивидуальных различиях темперамента см. в гл. X.) В дальнейших обследованиях ежегодно тестировались особенности умственного развития ребенка, некоторые показатели темперамента и личности, условия развития ребенка,
Масштабы этого исследования весьма внушительны, результаты еще полностью не опубликованы. Мы остановимся на некоторых
наиболее важных в данном контексте фрагментах. Речь в первую очередь идет об изучении генетически опосредуемой стабильности когнитивного развития. С целью определения роли генотипа в межвозрастной преемственности когнитивных характеристик анализировались межвозрастные кросс-корреляции сиблингов, т.е. подсчитывались корреляции между показателями одного сиблинга — младшего возраста и второго — более старшего возраста. Сравнение корреляций у биологических сиблингов, т.е. имеющих и общие гены и общую среду, а также приемных сиблингов, т.е. имеющих только общие средовые условия, позволили авторам определить вклад генотипа в межвозрастную стабильность и изменчивость когнитивных показателей (табл. 17.3). Анализ позволил выявить увеличение год от года роли генетических влияний в межвозрастной стабильности таких признаков, как общий интеллект и вербальные способности. Что же касается другого признака — пространственных способностей, то генетические влияния определяют его межвозрастную преемственность в более младших возрастах, но в 3-4 года решающую роль начинают играть средовые условия.
Таблица 17.3 Межвозрастные корреляции показателей интеллекта [по: 355]
| Показатели | Коэффициенты корреляции | |||||
| Родные сиблинги ЙПЧПЯГГ lYf. TTT-T*. | Приемные сиблинги ЙПЧПЯГГ lYf. TTT-T*. | |||||
| 1-2 | 2-3 | 3-4 | 1-2 | 2-3 | 3-4 | |
| Общий интеллект Способности: вербальные пространственные | 0221 | 18 18 | 09-11 | -03 -03 |
Сравнение IQ родителей и детей проводилось по трем вариантам: дети и их биологические родители, с которыми они были разлучены очень рано; дети и родители-усыновители; дети и биологические родители, с которыми они живут вместе, Результаты сравнения подтвердили значительную роль генетических факторов в опосредовании возрастной стабильности IQ.
В итоге можно констатировать, что вклад генотипа в индивидуальные различия интеллекта с возрастом увеличивается, причем генетические влияния в детском и взрослом возрасте преемственно связаны.
* * *
Соотношение генотипических и средовых влияний в формировании индивидуальных различий (в отличие от нормативного развития) является предметом многочисленных экспериментальных исследований в русле возрастной психогенетики (или психогенетики развития) — науки, изучающей природу межиндивидуальной изменчивости психологических особенностей человека в процессе онтогенеза. Генетические изменения в онтогенезе имеют два аспекта: изменения в сравнительной величине генетических и средовых компонентов межиндивидуальной изменчивости признака, т.е. изменения наследуемости; изменение генетической ковариации в ходе онтогенеза. В первом случае используется вариант метода возрастных срезов; во втором — лон-гитюдное исследование, в котором определяется корреляция между генетическими компонентами дисперсии изучаемой характеристики, полученными в разных возрастах на одной и той же группе испытуемых. Этот способ дает возможность оценить вклад генетических факторов в изменчивость возрастных преобразований,
Лонгитюдные исследования близнецов и приемных детей свидетельствуют о том, что вклад генотипа в индивидуальные различия IQ с возрастом увеличивается, причем генетические составляющие дисперсии интеллекта в младенчестве и в старших возрастах высоко коррелируют между собой. Это значит, что, несмотря на сравнительно низкий уровень наследуемости интеллекта в младенчестве, генетические эффекты, проявившиеся в столь раннем возрасте, продолжают оказывать влияние на интеллект человека и на более поздних этапах развития.
Методы психогенетики развития позволяют оценить возрастную динамику средовых эффектов. В детском возрасте средовая вариативность IQ определяется в основном действием факторов систематической семейной среды, т.е. общих для всех членов семьи, По мере взросления влияние систематической среды на IQ существенно снижается, но весьма ощутимо возрастает влияние уникальной, индивидуальной среды.
Глава XVIII
ВОЗРАСТНЫЕ АСПЕКТЫ ГЕНЕТИЧЕСКОЙ ПСИХОФИЗИОЛОГИИ
При анализе роли генотипа в формировании биоэлектрической активности мозга отмечалось, что в целом ряде случаев в исследованиях принимали участие близнецы разного возраста, причем возрастной разброс иногда оказывался весьма значительным. На первых по-
* * *
Соотношение генотипических и средовых влияний в формировании индивидуальных различий (в отличие от нормативного развития) является предметом многочисленных экспериментальных исследований в русле возрастной психогенетики (или психогенетики развития) — науки, изучающей природу межиндивидуальной изменчивости психологических особенностей человека в процессе онтогенеза. Генетические изменения в онтогенезе имеют два аспекта: изменения в сравнительной величине генетических и средовых компонентов межиндивидуальной изменчивости признака, т.е. изменения наследуемости; изменение генетической ковариации в ходе онтогенеза. В первом случае используется вариант метода возрастных срезов; во втором — лон-гитюдное исследование, в котором определяется корреляция между генетическими компонентами дисперсии изучаемой характеристики, полученными в разных возрастах на одной и той же группе испытуемых. Этот способ дает возможность оценить вклад генетических факторов в изменчивость возрастных преобразований,
Лонгитюдные исследования близнецов и приемных детей свидетельствуют о том, что вклад генотипа в индивидуальные различия IQ с возрастом увеличивается, причем генетические составляющие дисперсии интеллекта в младенчестве и в старших возрастах высоко коррелируют между собой. Это значит, что, несмотря на сравнительно низкий уровень наследуемости интеллекта в младенчестве, генетические эффекты, проявившиеся в столь раннем возрасте, продолжают оказывать влияние на интеллект человека и на более поздних этапах развития.
Методы психогенетики развития позволяют оценить возрастную динамику средовых эффектов. В детском возрасте средовая вариативность IQ определяется в основном действием факторов систематической семейной среды, т.е. общих для всех членов семьи, По мере взросления влияние систематической среды на IQ существенно снижается, но весьма ощутимо возрастает влияние уникальной, индивидуальной среды.
Глава XVIII
ВОЗРАСТНЫЕ АСПЕКТЫ ГЕНЕТИЧЕСКОЙ ПСИХОФИЗИОЛОГИИ
При анализе роли генотипа в формировании биоэлектрической активности мозга отмечалось, что в целом ряде случаев в исследованиях принимали участие близнецы разного возраста, причем возрастной разброс иногда оказывался весьма значительным. На первых по-
pax возрастным различиям не придавалось особого значения. Априори подразумевалось, что «генетическое» значит стабильное, неизменяющееся, поэтому возраст не является переменной, которую надо учитывать при оценке генетической детерминации психофизиологических признаков.
В настоящее время эта позиция уступила место другой: поскольку генетическая программа развития реализуется непрерывно, постольку признак, изменяясь в ходе развития, испытывает на себе влияния генотипа и среды, соотношения которых могут существенно меняться при переходе с одной стадии развития на другую. Такая логика хорошо согласуется с существующими в настоящее время представлениями о возрастной динамике созревания ЭЭГ и ВП.
1. ОСНОВНЫЕ ТЕНДЕНЦИИ В ФОРМИРОВАНИИ ЭЛЕКТРОЭНЦЕФАЛОГРАММЫ (ЭЭГ) И ВЫЗВАННЫХ ПОТЕНЦИАЛОВ (ВП)
Возрастные изменения биоэлектрической активности мозга охватывают значительный период онтогенеза от рождения до юношеского возраста. На основании многих наблюдений выделены признаки, по которым можно судить о зрелости биоэлектрической активности головного мозга. В их число входят: 1) особенности частотно-амплитудного спектра ЭЭГ; 2) наличие устойчивой ритмической активности; 3) средняя частота доминирующих волн; 4) особенности ЭЭГ в разных областях мозга; 5) особенности генерализованной и локальной вызванной активности мозга; 6) особенности пространственно-временной организации биопотенциалов мозга [173].
Наиболее изучены в этом плане возрастные изменения частотно-амплитудного спектра ЭЭГ в разных областях коры мозга. Для новорожденных характерна неритмичная активность с амплитудой около 20 мкВ и частотой 1-6 Гц. Первые признаки ритмической упорядоченности появляются в центральных зонах начиная с третьего месяца жизни. В течение первого года жизни наблюдается нарастание частоты и стабилизации основного ритма ЭЭГ ребенка. Тенденция к нарастанию доминирующей частоты сохраняется и на дальнейших стадиях развития. К 3 годам это уже ритм с частотой 7-8 Гц, к 6 годам — 9-10 Гц и т.д. [143]. Одно время считалось, что каждая частотная полоса ЭЭГ доминирует в онтогенезе последовательно одна за другой. По этой логике в формировании биоэлектрической активности мозга выделялись 4 периода: 1-й период (до 18 мес.) — доминирование дельта-активности, преимущественно в центрально-теменных отведениях; 2-й период (1,5 года — 5 лет) — доминирование тэта-активности; 3-Й период (6—10 лет) — доминирование альфа-активности (лабиль-
ная фаза); 4-й период (после 10 лет жизни) — доминирование альфа-активности (стабильная фаза). В двух последних периодах максимум активности приходится на затылочные области. Исходя из этого, было предложено рассматривать соотношение альфа и тэта-активности как показатель (индекс) зрелости мозга [430].
Однако проблема соотношения тэта- и альфа-ритмов в онтогенезе является предметом дискуссий. По одним представлениям, тэта-ритм рассматривается как функциональный предшественник альфа-ритма, и таким образом признается, что в ЭЭГ детей младшего возраста альфа-ритм фактически отсутствует. Придерживающиеся такой позиции исследователи считают недопустимым рассматривать доминирующую в ЭЭГ детей раннего возраста ритмическую активность как альфа-ритм [173]; сточки зрения других, ритмическая активность младенцев в диапазоне 6—8 Гц по своим функциональным свойствам является аналогом альфа-ритма [419].
В последние годы установлено, что альфа-диапазон неоднороден, и в нем, в зависимости от частоты, можно выделить ряд субкомпонентов, имеющих, по-видимому, разное функциональное значение. Существенным аргументом в пользу выделения узкополосных поддиапазонов альфа служит онтогенетическая динамика их созревания. Три поддиапазона включают: альфа-1 — 7,7-8,9 Гц; альфа-2 — 9,3-10,5 Гц; альфа-3 — 10,9-12,5 Гц. От 4 до 8 лет доминирует альфа-1, после 10 лет — альфа-2, и к 16-17 годам в спектре преобладает альфа-3 [143].
Исследования возрастной динамики ЭЭГ проводятся в состоянии покоя, в других функциональных состояниях (сои, активное бодрствование и др.), а также при действии разных стимулов (зрительных, слуховых, тактильных).
Изучение сенсорно-специфических реакций мозга на стимулы разных модальностей, т.е. ВП, показывает, что локальные ответы мозга в проекционных зонах коры регистрируются с момента рождения ребенка. Однако их конфигурация и параметры говорят о разной степени зрелости и несоответствия таковым у взрослого в разных модальностях [143]. Например, в проекционной зоне функционально более значимого и морфологически более зрелого к моменту рождения со-матосенсорного анализатора ВП содержат такие же компоненты, как и у взрослых, и их параметры достигают зрелости уже в первые недели жизни. В то же время значительно менее зрелы у новорожденных и младенцев зрительные и слуховые ВП.
Зрительный ВП новорожденных представляет собой позитивно-негативное колебание, регистрируемое в проекционной затылочной области. Наиболее значительные изменения конфигурации и параметров таких ВП происходят в первые два года жизни. За этот период ВП на вспышку преобразуются из позитивно-негативного колебания с латентностью 150-190 мс в многокомпонентную реакцию, которая в общих чертах сохраняется в дальнейшем онтогенезе. Окончательная стабилизация компонентного состава таких ВП
происходит к 5-6 годам, когда основные параметры всех компонентов зрительных ВП на вспышку находятся в тех же пределах, что и у взрослых. Возрастная динамика ВП на пространственно-структурированные стимулы (шахматные поля, решетки) отличается от ответов на вспышку. Окончательное оформление компонентного состава этих ВП происходит вплоть до 11-12 лет.
Эндогенные, или «когнитивные», компоненты ВП, отражающие обеспечение более сложных сторон познавательной деятельности, могут быть зарегистрированы у детей всех возрастов, начиная с младенчества [311], но в каждом возрасте они имеют свою специфику. Наиболее систематические факты получены при исследовании возрастных изменений компонента Р3 в ситуациях принятия решения. Установлено, что в возрастном диапазоне от 5-6 лет до взрослости происходит сокращение латентного периода и уменьшение амплитуды этого компонента. Как предполагается, непрерывный характер изменений указанных параметров обусловлен тем, что во всех возрастах действуют общие генераторы электрической активности.
Таким образом, исследование онтогенеза ВП открывает возможности для изучения природы возрастных изменений и преемственности в работе мозговых механизмов перцептивной деятельности.
ОНТОГЕНЕТИЧЕСКАЯ СТАБИЛЬНОСТЬ ПАРАМЕТРОВ ЭЭГ И ВП
Вариативность биоэлектрической активности мозга, как и другие индивидуальные черты, имеет две составляющих: внутрииндивиду-альную и межиндивидуальную. Внутрииндивидуальная вариативность характеризует воспроизводимость (ретестовую надежность) параметров ЭЭГ и ВП в повторных исследованиях. При соблюдении постоянства условий воспроизводимость ЭЭГ и ВП у взрослых достаточно высока. У детей воспроизводимость тех же параметров ниже, т.е. они отличаются значительно большей внутрииндивидуальной вариативностью ЭЭГ и ВП.
Индивидуальные различия между взрослыми испытуемыми (межиндивидуальная вариативность) отражают работу устойчивых нервных образований и в значительной степени определяются факторами генотипа. У детей межиндивидуальная вариативность обусловлена не только индивидуальными различиями в работе уже сложившихся нервных образований, но и индивидуальными различиями в темпах созревания ЦНС. Поэтому у детей она тесно связана с понятием онтогенетической стабильности. Это понятие подразумевает не отсутствие изменений в абсолютных значениях показателей созревания, а относительное постоянство темпа возрастных преобразований. Оценить степень онтогенетической стабильности того или иного показателя можно только в лонгитюдных исследованиях, в ходе которых сравниваются одни и те же показатели у одних и тех же детей на разных этапах онтогенеза. Свидетельством онтогенетической стабильно-
15-1432 385
сти признака может служить постоянство рангового места, которое занимает ребенок в группе при повторных обследованиях. Для оценки онтогенетической стабильности нередко используют коэффициент ранговой корреляции Спирмена, желательно с поправкой па возраст. Его величина говорит не о неизменности абсолютных значений того или иного признака, а о сохранении испытуемым своего рангового места в группе.
Таким образом, индивидуальные различия параметров ЭЭГ и ВП детей и подростков по сравнению с индивидуальными различиями взрослых имеют, условно говоря, «двойную» природу. Они отражают, во-первых, индивидуально устойчивые особенности работы нервных образований и, во-вторых, различия в темпах созревания мозгового субстрата и психофизиологических функций.
Экспериментальных данных, свидетельствующих об онтогенетической стабильности ЭЭГ, мало. Однако некоторые сведения об этом можно получить из работ, посвященных исследованию возрастных изменений ЭЭГ. В широко известной работе Линдсли [цит. по: 33] исследовались дети от 3 месяцев до 16 лет, причем ЭЭГ каждого ребенка прослеживалась в течение трех лет. Хотя стабильность индивидуальных особенностей специально не оценивалась, анализ данных позволяет заключить, что, несмотря на естественные возрастные изменения, ранговое место испытуемого примерно сохраняется.
Показано [33], что некоторые характеристики ЭЭГ оказываются устойчивыми в течение длительных периодов времени, невзирая на процесс созревания ЭЭГ. У одной и той же группы детей (13 чел.) дважды, с интервалом в 8 лет, регистрировалась ЭЭГ и ее изменения при ориентировочной и условно-рефлекторной реакциях в виде депрессии альфа-ритма. Во время первой регистрации средний возраст испытуемых в группе составлял 8,5 лет; во время второй — 16,5 лет, Коэффициенты ранговой корреляции для суммарных энергий составили: в полосах дельта- и тэта-ритмов — 0,59 и 0,56; в полосе альфа-ритма —0,36, в полосе бета-ритма —0,78. Аналогичные корреляции для частот оказались не ниже, однако наиболее высокая стабильность была выявлена для частоты альфа-ритма (R = 0,84).
У другой группы детей оценка онтогенетической стабильности таких же показателей фоновой ЭЭГ проводилась с перерывом 6 лет — в 15 лет и 21 год. В этом случае наиболее стабильными оказались суммарные энергий медленных ритмов (дельта- и тэта-) и альфа-ритма (коэффициенты корреляции для всех — около 0,6). По частоте максимальную стабильность вновь продемонстрировал альфа-ритм (Я = 0,47).
Таким образом, судя по коэффициентам ранговой корреляции между двумя рядами данных (1-е и 2-е обследования), полученным в этих исследованиях, можно констатировать, что такие параметры, как частота альфа-ритма, суммарные энергии дельта- и тэта-ритмов и ряд других показателей, ЭЭГ оказываются индивидуально стабильными.
Межиндивидуальная и внутрииндивидуальная вариативность ВП в онтогенезе изучена сравнительно мало. Однако один факт не вызывает сомнений: с возрастом вариабельность этих реакций уменьшает-
ся и нарастает индивидуальная специфичность конфигурации и параметров ВП [143]. Имеющиеся оценки ретестовой надежности амплитуд и латентных периодов зрительных ВП [107], эндогенного компонента Р3 [430], а также потенциалов мозга, связанных с движением [124], в общем говорят об относительно невысоком уровне воспроизводимости параметров этих реакций у детей по сравнению со взрослыми. Соответствующие коэффициенты корреляции варьируют в широком диапазоне, но не поднимаются выше 0,5-0,6. Данное обстоятельство существенно увеличивает ошибку измерения, которая, в свою очередь, может повлиять на результаты генетико-статистического анализа; как уже отмечалось, ошибка измерения включена в оценку индивидуальной среды. Тем не менее использование некоторых статистических приемов позволяет в таких случаях ввести необходимые поправки и повысить надежность результатов.
|
|
|
|
|
Дата добавления: 2014-11-20; Просмотров: 933; Нарушение авторских прав?; Мы поможем в написании вашей работы!