КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Представительство функций в коре большого мозга
|
|
|
|
Миелоархитектоника коры
В коре выделяют радиальные и тангенциальные нервные волокна. Первые вступают в кору из белого вещества или, наоборот, идут из коры в белое вещество полушария. Вторые располагаются параллельно поверхности коры и образуют в ней сплетения, называемые полосками. Различают полоски молекулярного слоя, наружного и внутреннего зернистого слоев и ганглионарного слоя. Количество полосок в различных частях коры неодинаково; в зависимости от этого описываются однополосковый, двухполосковый и многополосковый типы коры. Детальное исследование миелоархитектоники коры провели немецкие исследователи супруги Сесиль и Оскар Фогты, которые разделили полушарие более чем на 100 миелоархитектонических полей.
В центральной нервной системе с особенной полнотой проявляется диалектическое единство строения и функции. В свое время И. П. Павлов выдвигал в отношении нервной системы принцип структурности, согласно которому все мельчайшие детали конструкции мозга имеют динамическое значение. Исходя из этого, мы должны признать, что особенности архитектоники коры, различия в структуре областей и полей коры связаны с их функциональными отправлениями. Так мы подходим к вопросу о локализации, или представительстве, функций в коре большого мозга.
Учение о функциях мозга имеет многовековую историю. Значение мозга как органа ощущения и мысли понимали еще древнегреческие врачи Алкмеон и Гиппократ. В течение длительного времени свойство мышления приписывали «психической пневме» и локализовали психические функции в желудочках мозга. Леонардо да Винчи на одном из своих анатомических рисунков изобразил связь желудочков с opганом зрения посредством зрительного нерва, который рассматривался в то время как полая трубка, передающая зрительные впечатления в «передний» желудочек. Отсюда, как считал Леонардо да Винчи, ощущения передаются «общему чувству», расположенному «точно в середине головы». В XVII веке французский философ и ученый Р. Декарт считал, что в эпифизе мозга находится «седалище души», основываясь на том, что эпифиз лежит в центре полости черепа.
Первая попытка локализовать психические функции в определенных частях мозга;; была предпринята австрийским врачом Ф. И, Галлем, создавшим в конце XVIII века ученее -френологию. Согласно этому учению у человека имеется 37 душевных способностей, каждая из которых связана с особым участком полушария большого мозга. Величина и, форма полушарий отражают развитие тех или иных способ; костей л,.в свою очередь, отражаются на форме черепа. По: этому на основании осмотра и измерения головы можно судить о характере и способностях человека. Публичные выступления Галля носили материалистический и атеистический характер и были запрещены австрийскими властями, что вынудило ученого покинуть Вену. Учение Галля было вульгарно-материалистическим, и вскоре стала очевидной несостоятельность многих его положений, однако в нем содержалась правильная мысль о взаимосвязи между строением и функцией мозга.
Начало строго научному подходу к вопросу о локализации функций в коре положил французский ученый П. Брока, открывший в 1861 году двигательный центр речи в заднем отделе нижней лобной извилины левого полушария. В. А. Бец показал, что открытые им гигантские пирамидальные нейроны имеют отношение к двигательным центрам коры. В дальнейшем была определена локализация в коре различных видов чувствительности, выделены двигательные и ассоциативные зоны и стала преобладать тенденция к строгому пространственному делению коры в функциональном отношении.
Наряду с этим некоторые исследователи придерживались взгляда о функциональной равноценности, эквипотенциальности всех частей полушарий. Такой взгляд выдвигали в XIX веке французский физиолог Флуранс, производивший опыты на голубях, в XX веке американский психолог Лешли, ставивший эксперименты на крысах.
Как отмечает А. Р. Лурия, каждое из этих двух направлений внесло вклад в развитие учения о функциях мозга. Локализационистские взгляды способствовали разработке тонкой анатомии мозга и представлений о дифференцированной работе его частей. Сторонники эквипотенциализма развивали положения о высокой пластичности мозговой ткани и способности к восстановлению функций, нарушенных в результате ее повреждения.
Большую роль в изучении локализации функций в коре большого мозга сыграли работы И. П. Павлова и сто школы. И. П. Павловым была заложена основа динамического подхода к локализации функций. Согласно учению И. П. Павлова нервная система представляет комплекс анализаторов внешней и внутренней среды организма. В коре большого мозга находятся центральные концы анализаторов, которые осуществляют анализ и синтез сигналов, поступающих к ним по афферентным нервным путям.. На основании экспериментов на собаках И.П. Павлов установил, что корковые концы анализаторов не имеют резко очерченных границ и что и х периферические части заходят друг за друга, образуя обширные зоны перекрытия.
За последние десятилетия в изучении функций коры широко применялись электрофизиологические методики, а также наблюдения на больных людях при нейрохирургических операциях. В настоящее время признается, что различные • корковые поля являются функционально многозначными, то есть участвуют в нескольких функциях, выступая в каждой из них в сочетаниях с другими мозговыми структурами. Поэтому любая функция коры обычно является результатом деятельности ряда взаимосвязанных ее» частей. Архитектоника коры составляет анатомическую о снову понимания ее сложной, системной деятельности.
В структурно-функциональном отношении кора большого мозга делится на передний и задний отделы. Граница между ними проходит по центральной борозде. Таким образом, задний отдел полушария образован теменной, затылочной и височной долями, а передний отдел представлен лобной долей. Задний отдел коры связан с восприятием и переработкой афферентных сигналов, то есть представляет комплекс корковых концов анализаторов. Передний отдел коры имеет отношение к организации действий.
Принято подразделять цитоархитектонические поля коры на первичные, вторичные и третичные (рис. 1).
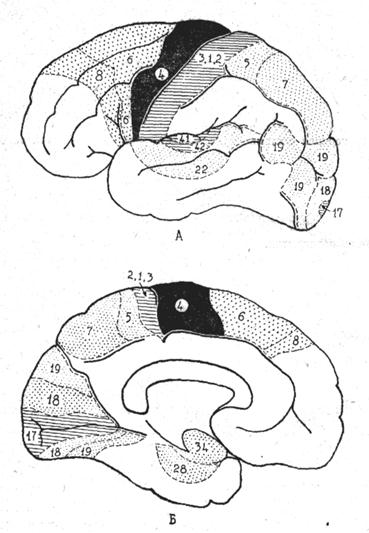
|
Рис. 9. Локализация первичных и вторичных корковых полей. Первичные чувствительные поля заштрихованы, первичное двигательное поле черное. Вторичные чувствительные поля выделены мелким, вторичные двигательные поля---крупным пунктиром. Третичные поля коры оставлены белыми. Нумерация корковых полей по Бродману.
Первичные поля представляют четко локализованные участки, которые соответствуют центральным (ядерным) частям анализаторов. В эти поля по специфическим афферентным путям приходит основная масса сигналов от органов чувств. В архитектоническом отношении первичные поля характеризуются сильным развитием внутреннего зернистого слоя, в котором, как мы знаем, оканчивается большинство афферентных волокон. Первичные поля связаны с реле-ядрами таламуса и коленчатых тел. Они имеют экранную структуру и, как правило, пространственную проекцию периферии. Повреждение первичных воспринимающих нолей приводит к нарушению непосредственного восприятия и топкой дифференцировки раздражений.
Представительство кожной и сознательной проприоцептивной чувствительности занимает первичные корковые поля 1, 2 и 3, расположенные в постцентральной извилине. В каждом полушарии имеется обратная проекция противоположной половины тела. В верхней части извилины находится проекция нижней конечности, ниже располагается проекция живота, груди, верхней конечности. В нижней трети извилины локализуются раздражения, приходящие от головы и шеи, причем проекция частей головы и шеи является уже прямой. Первичное зрительное поле (17) расположено на медиальной поверхности полушария вдоль шпорной борозды; над бороздой проецируются верхние, а под бороздой нижние квадранты сетчаток. В первичном зрительном поле хорошо развит внутренний зернистый слой коры, который подразделяется на 3 части посредством волокнистых белых полосок (area striata).
Первичные слуховые поля (41 и 42) локализуются в поперечных височных извилинах и заходят в верхнюю височную извилину.
Первичная обонятельная область захватывает древнюю кору, покрывающую обонятельный треугольник, переднее продырявленное пространство, прозрачную перегородку и подмозолистое поле.
Первичное вкусовое поле располагается, по-видимому, в нижнем участке постцентральной извилины, в глубине латеральной борозды. По другим данным, оно находится в крючке.
Точно не установлено нахождение поля, принимающего раздражения от вестибулярного аппарата.
Вторичные поля коры примыкают к первичным, их можно рассматривать как периферические части корковых концов анализаторов. Вторичные поля связаны с ассоциативными ядрами таламуса, в них преобладают нейроны с короткими отростками, за счет которых происходит взаимодействие сигналов, поступающих по различным афферентным системам и из первичных полей. При поражении вторичных полей сохраняются элементарные ощущения, однако нарушается способность к более сложным восприятиям.
Вторичные поля не имеют строго очерченных границ, и в них отсутствует пространственная проекция периферии. Вторичное поле общей чувствительности располагается в верхней теменной дольке (поля 5 и 7). Вторичные зрительные поля занимают медиальную и большую часть латеральной по-верхности затылочной доли (поля 18 и 19). Вторичное слуховое поле 22 находится в верхней и средней височных извилинах. Вторичные обонятельные и вкусовые поля локализуются в парагиппокампальной извилине и крючке (поля 28 и 34).
Представительство в коре внутренностей и сосудов изучено недостаточно, по-видимому оно имеет более диффузный характер, чем представительство покрова тела и мускулатуры. Важная роль в регуляции функций внутренних органов придается лимбической области коры, в которую входят поясная и парагиппокампальная извилины, гиппокамп, перегородка и подмозолистое поле. Лимбическая кора вместе с миндалевидным телом и некоторыми другими подкорковыми образованиями составляют лимбическую систему, которая представляет субстрат эмоций и реакций, связанных я основными биологическими, влечениями (голод, жажда,страх,- ярость, половое возбуждение).
Третичные поля коры отличаются, наиболее тонкой нейронной структурой с преобладанием ассоциативных элементов: Третичные поля занимают в заднем отделе полушария всю нижнюю и часть верхней теменной долек и затылочно-височно-тменную область. Они связаны с задним ядром (подушкой) таламуса. В третичных полях осуществляются наиболее сложные формы взаимодействия анализаторов, лежащие в основе познавательного процесса (гнозия), формируются программы целенаправленных действий (праксия).
Кора височной доли имеет отношение к хранению и воспроизведению впечатлений. Известный канадский учёный; нейрохирург Уилдер Пенфилд установил, что при электрическом раздражении некоторых участков височной коры возникают своеобразные реакции в форме «вспышек пережитого» или ощущения «уже виденного». При этом переживается как действительность какой-то отрезок прошлого или больному кажется, что операция уже производилась раньше и он знает, что собирается делать хирург. Пенфилд предполагает, что в коре височных долей создается нейронная запись потока сознания, она хранится неопределенно долгое время, но человек не может произвольно воспроизводить эту «запись», и, она активируется лишь при искусственной симуляции или болезненных состояниях.
Передний отдел полушария также подразделится на: первичные, вторичные и третичные корковые поля. Первичное двигательное поле располагается в предцентральной извилине (поле 4). Здесь отсутствует внутренний зернистый слой (агранулярная кора) и особенно сильно развит ганглионарный слой, в котором находятся гигантские пирамидальные нейроны. Аксоны этих клеток образуют пирамидный путь. На гигантские пирамидальные нейроны непосредственно переключаются импсульсы, приходящие сюда по афферентным волокнам из мозжечка через центральное медиальное ядро таламуса. В первичном двигательном поле представлены все мышечные группы тела. Как и в первичном чувствительном поле, здесь имеется обратная проекция частей тела. Верхние участки предцентральной извилины связаны с мускулатурой нижних конечностей, далее книзу находится проекция мускулатуры туловища, верхней конечности/головы и шеи. Первичное двигательное поле регулирует индивидуальные сокращения мышц, от которых зависит выполнение произвольных движений.
Вторичным двигательным полем является кора лобных извилин кпереди от предцентральной извилины (поля 6 и 8). Они характеризуются сильным развитием пирамидного слоя, в котором преобладают крупные пирамидальные нейроциты. Они, как и гигантские клетки первичного двигательного поля, получают импульсы из мозжечка. Вторичное двигательное поле является корковым центром экстрапирамидной системы; эфферентные волокна идут из него в полосатое тело.
Сигналы, посылаемые вторичным двигательным полем, необходимы для осуществления сложных стереотипных двигательных актов.
Поля 4 и 6 имеют многочисленные связи с задним отделом полушария. Роль двигательной коры состоит в том, что в ней сопоставляются программы движений с реальным ходом выполнения этих программ и вносятся коррекции по ходу выполняемых действий. Обратная связь между двигательным аппаратом и корой осуществляется через мозжечок, который воспринимает афферентные импульсы с периферии и после соответствующей переработки сигналов направляет их в кору.
Третичные поля занимают большую часть поверхности лобной доли и четверть всей коры большого мозга. В них хорошо выражен внутренний зернистый слой, в который приходят импульсы из медиальных ядер таламуса. Третичные поля лобной коры имеют отношение к высшим формам целенаправленной деятельности и социальному поведению человека. При их поражении не возникает каких-либо чувствительных или двигательных расстройств, но человек становится пассивным, безучастным к происходящему, теряет способность к оценке событий и предвидению.
Важнейшую особенность человека составляет членораздельная речь. И. П. Павлов относил речь ко второй сигнальной системе, с помощью которой осуществляется непрямое отражение действительности. Речевые функции имеют широкое представительство в коре большого мозга. И. П. Павлов полагал, что двигательная речь должна быть связана с двигательной областью коры, письменная речь — со зрительными областями, а слышимая речь — с височными долями. У. Пенфилд и Л. Роберте, основываясь на данных, полученных при электрическом раздражении и удалении у больных различных участков коры, выделили в коре три речевых поля (рис. 2).
Заднее речевое поле располагается в затылочно-височно-теменной области, захватывая все три височные, надкраевую и угловую извилины. Это поле связано преимущественно с восприятием и пониманием речи и в функциональном отношении является ведущим. При его поражении всегда наступает афазия — расстройство речи.
Переднее речевое поле находится в задней части нижней лобной извилины и соответствует двигательному центру речи Брока.
Верхнее, дополнительное речевое поле локализуется у верхнего края полушария и заходит на его медиальную поверхность кпереди от парацентральной дольки. Поражение этого участка не всегда вызывает речевые расстройства.
Речевые поля, как и другие части коры, связаны с ядрами таламуса. Заднее поле связано с задним ядром, верхнее поле— с латеральным ядром, переднее поле — с медиальными ядрами таламуса.
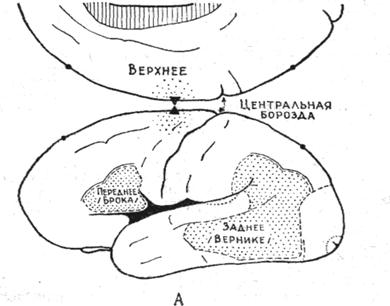
Рис. 10. Речевые поля коры по Пенфилду и Робертсу.
Все речевые поля объединяются посредством нервных связей, образуя единую функциональную систему.
Замечательной особенностью речевых центров коры является их асимметрия. У большинства людей они располагаются в левом полушарии, которое является доминантным г. отношении речи. Принято считать, что доминантность левого полушария связана с праворукостью и что у левшей речевые функции локализуются в правом полушарии. В противоположном субдоминантном полушарии соответствующие участки коры выполняют иные функции: с ними связано представление о пространственных отношениях и схеме тела.
|
|
|
|
|
Дата добавления: 2014-12-10; Просмотров: 1518; Нарушение авторских прав?; Мы поможем в написании вашей работы!