КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пассивный транспорт веществ и его разновидности. Математическое описание пассивного транспорта
|
|
|
|
Пассивный транспорт веществ через клеточные мембраны включает в себя следующие разновидности: простую диффузию и облегченную диффузию Фильтрация
Простая диффузия – процесс переноса вещества из области с большей концентрацией в область меньшей концентрации. Диффузия описывается уравнением Фика:
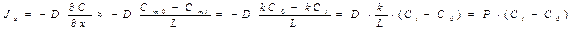 ,
,
где 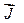 вектор плотности потока массы диффундирующего вещества,Сi и C0 – концентрации диффундирующего вещества внутри и вне клетки. Сmi и Cm0 – концентрации диффундирующего вещества внутри мембраны на границе мембрана.P– проницаемость мембраны.
вектор плотности потока массы диффундирующего вещества,Сi и C0 – концентрации диффундирующего вещества внутри и вне клетки. Сmi и Cm0 – концентрации диффундирующего вещества внутри мембраны на границе мембрана.P– проницаемость мембраны.
Облегченная диффузия.
Срособность мембран обусловлена двумя причинами: наличием в них переносчиков, называемых ионофорами (подвижных и фиксированных) и каналов. Облегченная диффузия – процесс транспорта веществ с помощью специальных молекул-переносчиков.
Фильтрация. Фильтрация представляет собой перенос молекул растворителя под действием градиента давления. Таким образом, причиной и движущей силой в процессе фильтрации является разность давлений. Объем растворителя, перенесенного в результате фильтрации, определяется формулой Пуазейля:
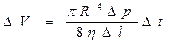 ,
,
где 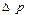 - перепад давления на расстоянии
- перепад давления на расстоянии 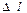 ,
, 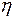 - вязкость жидкости,
- вязкость жидкости, 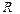 - радиус поверхности фильтрации,
- радиус поверхности фильтрации, 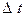 - время переноса.
- время переноса.
Фильтрация. Фильтрация представляет собой перенос молекул растворителя под действием градиента давления. Таким образом, причиной и движущей силой в процессе фильтрации является разность давлений. Объем растворителя, перенесенного в результате фильтрации, определяется формулой Пуазейля:
,
где - перепад давления на расстоянии , - вязкость жидкости, - радиус поверхности
жидкости.
54. Активный транспорт ионов. Механизм активного транспорта вещ-в на примере Na-K насоса. Если бы в клетках сущ. только пассивный транспорт, то конц-ции, давления и др. величины вне и внутри клетки сравнялись бы. Поэтому сущ-т др. механизм, работающий в направлении против электрохимического градиента и происходящий с затратой энергии клеткой. Перенос молекул и ионов против электрохимического градиента,осуществляемый клеткой за счет энергии метаболических процессов, наз. активным транспортом. Он присущ только биологическим мембранам. Активный перенос вещ-ва через мембрану происходит за счет свободной энергии,высвобождающейся в ходе хим. Реакций внутри клетки.Активный транспорт в организме создает градиенты концентраций, электр. потенциалов, давлений,т.е. поддерживает жизнь в организме. Изучены 3 основные системы акт.трансп., кот-ые обеспечивают перенос ионов Na,K,Ca,H через мембрану. Механизм. Ионы К+ и Na+ неравномерно распределены по разные стороны мембраны:концентр. Na+ снаружи > ионов K+,а внутри клетки K+ > Na+.Эти ионы диффундируют через мембрану по направлению электрохимического градиента,что приводит к его выравниванию. Na-K насосы вх. в состав цитоплазмат. мембран и работают за счет энергии гидролиза молекул АТФ с обр-ем мол-л АДФ и неорганич. фосфата Фн: АТФ=АДФ+Фн. Насос работает обра-тимо: градиенты конц-ций ионов способ-ют синтезу мол-л АТФ из мол-л АДФ и Фн: АДФ+Фн=АТФ.Насос переносит из клетки во внеш. среду 3 иона К+внутрь клетки.
55. Способы проникновения вещ-в через биологические мембраны.. Одной из важнейших хар-к клеточных мембран(КМ) явл-ся избират. проницаемость. КМ избирательно снижает скорость передвижения мол-л в клетку и из нее. Чем меньше мол-ла и чем меньше она обр. водород. связей,тем быстрее она диффун-дирует ч/з мембрану. =>, чем меньше мол-ла и чем более она жирорастворима, тем быстрее она будет проникать через мембрану.Малые неполярные мол-лы легко растворимы в липидах КМ и быстро диффундируют.Клетка была вынуждена создать спец.мех-мы для транспорта растворимых в воде вещ-в через мембрану-через поры в мембране и посредством транспортных белков-переносчиков мол-л. Для жиронерастворимых вещ-в и ионов мембрана выступает как молекулярное сито: чем больше размер частицы,тем меньше проницаемость мембраны для этого вещ-ва. Избирательность переноса обеспечивается набором в мембране пор определенного радиуса,соответствующих размеру проникающей частицы.Это распределение зависит от мембранного потенциала. Перенос малых водорастворимых мол-л осуществляется при помощи специальных транспортных белков.Это особые белки,каждый из которых отвечает за транспорт определенных мол-л или групп мол-л. За перенос сахара,аминокислот и др. полярных мол-л ответственны специальные мембранные транспортные белки.Каждый из них предназначен для определенного класса мол-л.Все они обеспечивают перенос мол-л через мембрану, формируя в ней сквозные проходы. Транспортные белки делятся набелки-переносчики, и каналообразующие белки. Переносчики взаим-ют с молекулой переносимого вещ-ва и каким-либо способом перемещают ее сквозь мембрану. Каналообразующие -формируют в мембране водные поры,через кот-ые могут проходить вещ-ва. Отличия облегченной диффузии от простой: 1)перенос ионов с участием переносчиков происх. значительно быстрее; 2)обладает св-вом насыщения- при ув. концентр. С одной стороны мемраны плотность потока вещ-ва возрастает лишь до некоторого предела. Разновидностью облегч.дифф.-транспорт с помощью неподвижных мол-л переносчиков,фиксированных поперек мембраны. Осмос- движение мол-л воды через полупроницаемые мембраны из мест с меньшей концентрацией растворенного вещ-ва в места с большей концентр. Осмос обусл-ет гемолиз эритроцитов в гипотонических растворах и тургор в растениях.
56. История открытия биопотенциалов. Гипотеза Бернштейна.. Фр.священник аббат Нолле в 1746г открыл явление осмоса.1826г. Дютроше доказал,что осмос есть результат проявления не особых,мифических сил,а законов физики и химии. Немецк.ботаник Пфеффер -изобрел осмометр и измерил величину осмотического давления.Он обнаружил,что для каждого раствора величина давления пряма-пропорциональна концентр. раств.вещ-ва,не проходящего через полупроницаемую мембрану (Росм=m/V m-масса растворенного вещ-ва, V-объем раствора). Вант-Гофф пришел к заключению,что мол-лы растворенного вещ-ва в растворителе ведут себя подобно мол-ам идеального газа(Росм=СмRT См-молярная плотность растворенного вещ-ва).Теория Вант-Гоффа давала точные значения величины осм.давления для многих вещ-в,но для некоторых оказ. больше расчетной в 2раза. С.Аррениус предположил что в р-ре мол-ла соли распадается на 2 частицы-электролитич. диссоциация. Вальтер Нернст обосновал идею диффузного потенциала, возн-го при соприк-нии двух жидкостей. Вел-на дифф.потенц. рассчит-ся: Фн=(u-v/u+v)×(RT/F)×Ln(C1/C2), где u и v -скорости быстрого и медл. ионов, R- газ.постоянная , C1 и С2- конц-ции электролита. Для возн-ния дифф.потенц. нужна разность конц-ций электролита, различная подвиж-ть анионов и катионов. Бернштейн начал объяснять электрич св-ва мышц не устройством этих органов в целом,а свойствами клето,из которых эти органы состояли. 1902г-год рождения мембранной теории биопотенциалов. Согласно гипотезе Бернштейна, каждая клетка им. оболочку,кот-ая представляет собой полупроницаемую мембрану. Внутри и вне клетки имеется много свободных ионов,среди кот-ых нах-ся ионы K+. Разность пот-лов между внутр. стороной БМ и ее наружной стороной наз-ся потенциалом покоя (ПП).Величина ПП опис-ся формулой Нернста: ФН=-(RT/F)×Ln[(K+)I/(K+)0] где [K+]i- конц-ция ионов К внутри клетки, [K+]0- конц-ция ионов К снаружи клетки.
57. Мембранно-ионная теория генерации биопот-лов клеткой и основ. опыты, ее подтвер-щие. Ю.Бернштейн (опыты на мышце лягушки). Нагрев 1 конца целой мышцы, от нагретого участка к холодному потечет ток. Электрич. ток течет по направлению от точек пространства с более высоким потенциалом к месту с более низким значением электрического потенциала.В 1905г. Гебер обнаружил,что все соли,сод-е К, оказ. на мышцу схожее д-вие: участок на кот-ый действовал раствор соли К, приобретал отрицательный потенциал по отношению к другим участкам мышцы.Все соли К при диссоциации в воде повышали наружную конц-цию ионов К, при этом отношение (K+)i\(K+)0 умен-ся, ум-ся и пот-ал той области мышцы, на кот. д-ют соли К. Однако эксперименты Бернштейна и Гебера были косвенными. Чтобы подтвердить правильность гипотезы,требовалось доказать следующее: 1)клетки им. мембрану,кот-ая проницаема лишь для одного иона;2)конц-ция этого иона по обе стороны БМ различная;3)потенциал на мембране возникает только за счет проницаемости мембраны для этого иона и он равен нернстовскому потенциалу. В 1936г. Дж.Юнг обнаружил кольмара,у кот-ого диаметр нервного волокна доходил до миллиметра.Аксон кальмара был гигантской клеткой,хотя сам моллюск не был гигантом.Нервное волокно выняли из моллюска и поместили в морскую воду,и оно не погибло.=>(эксперимент на клет.уровне). В 1939г. А.Ходжкин и Хаксли измерили разность потенциалов на аксоне кальмара.Они доказали,что внутри аксона им. много ионов К,и они обр. ионный газ,т.е.нах-ся в свобод. сост-нии.
|
|
|
|
|
Дата добавления: 2014-12-16; Просмотров: 1597; Нарушение авторских прав?; Мы поможем в написании вашей работы!