КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Фоторецепторы
|
|
|
|
Структуры молекул и ячеек в рецепторных клетках для семи основных запахов
Обоняние.
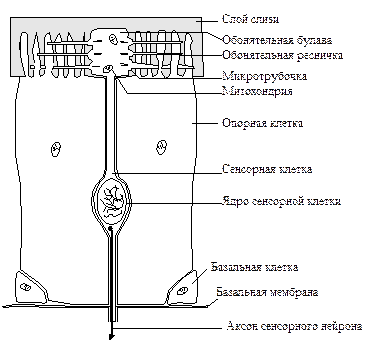
[рисунок обонятельного центра]
Одно время считали, что в основе всего разнообразия запахов лежат всего 7 «основных» запахов:
– камфорный
– мускусный
– цветочный
– мятный
– эфирный
– едкий
– гнилостный
| Камфорный | Мускусный | Цветочный |
| 
| 
|

| 
| 
|
| 
| 
|
| Мятный | Эфирный | Едкий |
| 
| 
|

| 
| 
|
| 
| Гнилостный


|
Была высказана гипотеза, что обонятельные рецепторы отличаются своей формой, специфичной для каждого «основного» запаха. Эта гипотеза оказалась неверной.
С помощью глаз мы получаем до 90% информации об окружающем мире. Глаз способен различать свет, цвет, движение, способен оцениать скорость передвижения. Максимальная концентрация светочувствительных клеток глаза там, где оптическая ось (рассматриваем глаз как оптическую систему) пересекается с дном глаза.
Радужная оболочка играет роль диафрагмы, регулируя количество света, проникающего в глаз. Она очень сильно иннервирована, причем на ней присутствуют клетки, комплементарные многим органам. На радужку вынесен как бы «портрет» организма. Существует даже методика диагностики различных заболеваний только по радужке.
Функционирование.
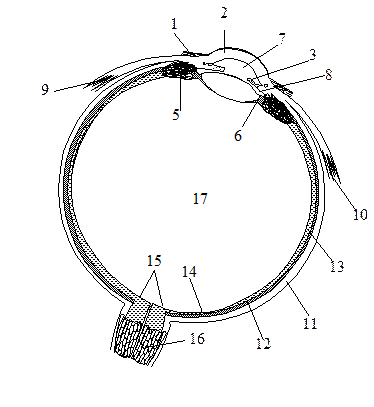
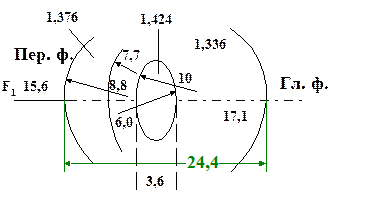
Рис. Горизонтальный разрез глазного яблока (схема).
1 – конъюнктива; 2 – роговица; 3 – радужка; 4 – хрусталик; 5 – ресничное тело; 6 – связка, при помощи которой хрусталик прикрепляется к ресничному телу (ресничный поясок); 7 – передняя камера глаза; 8 – задняя камера глаза; 9,10 – мышца глазного яблока; 11 – склера; 12 – собственно сосудистая оболочка; 13 – сетчатка; 14 – центральная ямка; 15 – решетчатая пластинка; 16 – зрительный нерв; 17 – стекловидное тело.
Существует несколько заблуждений относительно функционирования глаза. Первое из них состоит в том, что основная оптическая сила глаза формируется хрусталиком. Это не так. Основная оптическая сила формируется поверхностью роговицы. Роговица имеет две поверхности с разными радиусами кривизны. Оптическая сила глаза варьируется от 70,6 до 58,6 дптр. в 10 лет 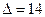 дптр, а в 70 лет
дптр, а в 70 лет 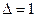 дптр. с возрастом хрусталлик теряет свою выпуклость, и растягивающие мышцы его пересиливают, делая его ещё более плоским.
дптр. с возрастом хрусталлик теряет свою выпуклость, и растягивающие мышцы его пересиливают, делая его ещё более плоским.
| 
| ||||
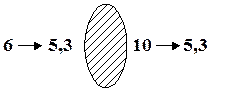
| 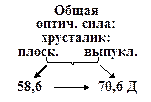
| 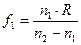
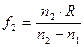
| |||
| 10 лет | D = 14 Д | ||||
| 70 лет | D = 1 Д | ||||
| 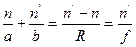
| ||||
| Дрейф 2¸10’ 6’ / сек скачок неск. 0/сек! | 
| ||||
Существует понятие «приведенного глаза»: если из кварца вырезать фигуру, по форме напоминающую глаз, выдержав кривизну поверхностей, то такая система, подобно глазу будет фокусировать свет.
Второе заблуждение состоит в том, что глаз является тонкой линзой. На самом деле глаз является толстой линзой, для которой можно написать соответствующие формулы.
Глаз никогда не стоит на месте. Зрачок совершает постоянный дрейф со скоростью 2’ – 10’/сек. Возможно это связано с тем, что если зафиксировать источник света так, чтобы он фокусировался на одном участке сетчатки, то через секунду его изображение исчезнет.
Существует понятие остроты зрения (собственно, критерий Рэлея).
Острота:
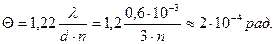
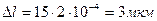
У сетчатки выделяют 2 слоя: верхний и глубинный. Верхний составляют кровеносные сосуды и нервные клетки, а глубинный – светочувствительные клетки и пигментные клетки (так называемый «черный эпителий»). Роль этих пигментных клеток остается невыясненной. Раньше думали, что он нужен только для того, чтобы поглощать «лишний» свет, однако при различной освещенности этот эпителий изменяет форму (?). у кошек этот эпителий в темноте становится просто отражающим экраном, это объясняет то, что у кошек в темноте глаза «светятся». Т.о. прежде чем свет попадет на светочувствительные клетки, он проходит побочные, рассеивающие ткани, в которых теряется до 50% попадающего в зрачок света.
Светочувствительные клетки глаза бывают двух видов: колбочки и палочки. Палочек примерно 120 млн., колбочек – 6 млн. плотность колбочек максимальна в середине центральной ямки, а палочек – вокруг этой ямки; в ней самой они полностью отсутствуют. Оба типа рецепторов сходны по структуре в том смысле, что и у тех, и у других есть наружный сегмент, состоящий примерно из тысячи мембранных дисков (палочки) и мембранных складок (колбочки). Он соединяется с остальной частью клетки узкой «ресничкой». Время жизни дисков равен приблизительно 10 дней. Палочки представляют собой вторично-чувствующие рецепторы.
На первичные рецепторы попадает сигнал (возбуждение) от нескольких колбочек. В процессе передачи сигнала на более «поздние» нейроны изменяются характеристики электрических сигналов так, что если палочки генерируют аналоговый сигнал, то с нервных клеток можно снять последовательность прямоугольных импульсов.
 Латеральное торможение – торможение сигналов соседних клеток за счет своего возбуждения. Благодаря этому подчеркивается граница между светлым и темным. Это можно видеть на графике. Интенсивность сигналов «теневых» клеток (тех, на которые попадает мало света) уменьшается соседними возбужденными (теми, на которые попадает много света) клетками. Этим объясняется провал. Возбужденные клетки слабо тормозятся соседними «теневыми» ® на графике виден «горб».
Латеральное торможение – торможение сигналов соседних клеток за счет своего возбуждения. Благодаря этому подчеркивается граница между светлым и темным. Это можно видеть на графике. Интенсивность сигналов «теневых» клеток (тех, на которые попадает мало света) уменьшается соседними возбужденными (теми, на которые попадает много света) клетками. Этим объясняется провал. Возбужденные клетки слабо тормозятся соседними «теневыми» ® на графике виден «горб».
 |
Такая первичная обработка сигналов происходит без участия мозга, в этом смысле глаз и сетчатку называют мозгом, вынесенным наружу.
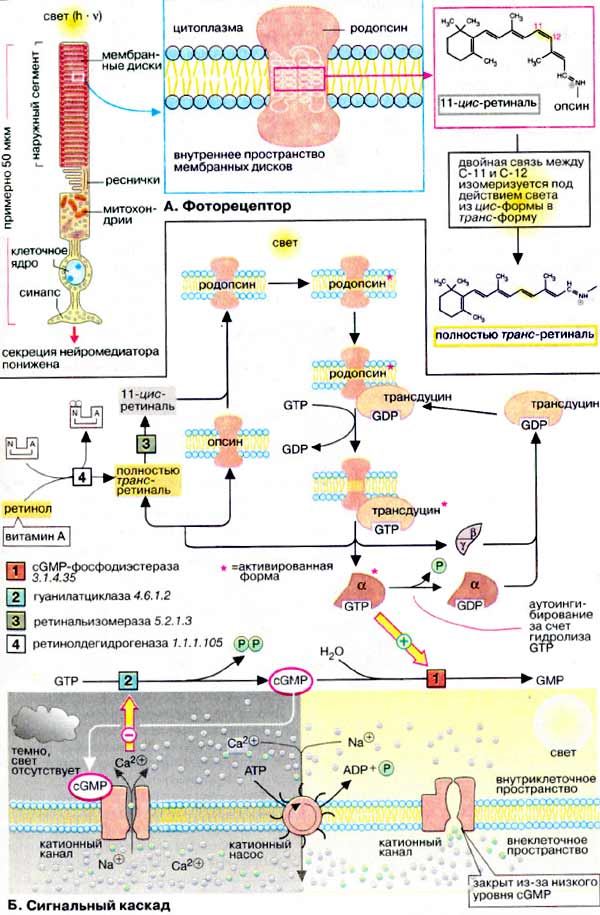
В сетчатке глаза позвоночных содержатся два типа фоторецепторных клеток: палочки и колбочки. Палочки чувствительны к свету, а колбочки отвечают за восприятие цвета.
А. Фоторецептор
На рисунке схематически изображена одна из фоторецепторных клеток, палочка. Клетка состоит из двух основных частей, наружного и внутреннего сегментов. В дисках наружного сегмента (специализированных замкнутых мембранах) локализован родопсин, интегральный мембранный белок, включающий 7 трансмембранных тяжей. Такое строение характерно для большой группы сигналпереносящих рецепторных белков. Родопсин является светочувствительным хромопротеином. Помимо белковой части, опсина, молекула родопсина включает остаток 11-цис-ретиналя, связанный ковалентно с ε-аминогруппой остатка лизина. Родопсин обладает характерным спектром поглощения света с максимумом при 500 нм.
Поглощение молекулой родопсина кванта света индуцирует изомеризацию 11-цис-ретиналя в полностью транс-форму. В результате этой фотохимической реакции изменяется геометрия ретиналя, а спустя 10 мс происходит аллостерический переход родопсина в его активную форму (родопсин*). Стимуляция родопсином* G-белка запускает каскад передачи сигнала, который побуждает зрительную клетку уменьшить выброс нейромедиатора (глутамата), вследствие чего биполярные нейроны, связанные со зрительными клетками, посылают измененный импульс, что воспринимается как зрительное возбуждение.
Б. Сигнальный каскад
G-белок палочек носит название трансдуцин. Связывание активированного светом родопсина* (метародопсина II) с ГДФ-трансдуцином катализирует обмен ГДФ (GDP) на ГТФ (GTP). Активная форма трансдуцина (ГТФ-трансдуцин) диссоциирует на комплекс β, γ-субъединиц и ГТФ-α*-субъединицу (см. с. 372), которая активирует цГМФ-фосфодиэстеразу (сGΜΡ - фосфодиэстеразу) [1], связывая ингибиторную субъединицу фермента.
В отсутствие света концентрация цГМФ (cGMP) в колбочках поддерживается на сравнительно высоком уровне (70 мкМ). Этот вторичный мессенджер (см. с. 374) постоянно синтезируется гуанилатциклазой и гидролизуется цГМФ-фосфодиэстеразой. Активация фосфодиэстеразы (при освещении родопсина) вызывает быстрое (в течение нескольких мс) падение уровня цГМФ.
Спустя короткое время α-субьединица трансдуцина инактивируется за счет медленного гидролиза связанного ГТФ и ассоциирует с комплексом β, γ-субъединиц. Родопсин* распадается на опсин и полностью транс-ретиналь, который изомеризуется в цис-ретиналь под действием изомеразы [3]. После сборки родопсина молекула возвращается в исходное состояние.
В темноте (на схеме слева внизу) высокий уровень цГМФ в палочках поддерживается благодаря активности гуанилатциклазы. Поэтому цГМФ-зависимые катионные каналы плазматической мембраны остаются открытыми и катионы Na+ и Ca2+ беспрепятственно поступают в клетку. При этом зрительная клетка постоянно выбрасывает нейромедиатор глутамат в синаптическую щель.
При освещении (на схеме справа внизу) уровень цГМФ резко падает за счет активации фосфодиэстеразы*, что приводит к перекрыванию ионных каналов. Так как ионы Na+ и Ca2+ постоянно выкачиваются из клетки, концентрация их быстро падает. Это приводит к гиперполяризации клетки и останавливает выброс нейромедиатора. Снижение концентрации ионов Ca2+ инициирует активацию гуанилатциклазы, что влечет за собой быстрый подъем уровня цГМФ настолько, что ионные каналы открываются вновь.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 659; Нарушение авторских прав?; Мы поможем в написании вашей работы!