КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Антитела
|
|
|
|
Антителам присущи следующие основные биологические
свойства.
1. Специфичность — способность Ig реагировать только с определенным антигеном, что обусловлено наличием у них антидетерминант, контактирующих с соответствующими детерминантами.
2 Валентность — количество антидетерминант в молекуле антитела. Чаще всего антитела бивалентны, но существуют 5- и даже 10-валентные.
3. Аффинность, аффинитет — прочность соединения между детерминантами (эпитопом) антигена и антидетерминантами (пара-топом) антигена.
4 Авидность — прочность связи антигена с антителом, ълаго-даря поливалентности антигена связь между двумя антигенами осуществляется при помощи нескольких антител.
5. Гетерогенность — неоднородность, обусловленная наличием трех видов антигенных детерминант:
A. Изотипические, характеризующие принадлежность Ig к оп
ределенному классу;
Б. Аллотипические, соответствующие аллельным вариантам иммуноглобулина;
B. Идиотипические, отражающие индивидуальные особеннос
ти иммуноглобулина.
Организм способен реагировать на поступление чужеродных антигенов синтезом белков, обладающих специфическим срод-
ством с антигеном, вызвавшим этот синтез. Эти белки получили название антител. Хотя антитела отличаются друг от друга не только по способности связывать разные антигены, но и по своим физико-химическим свойствам, структурно все они сходны. Эта высокоспециализированная группа белков, построенных из поли-пептидных цепей двух основных типов (легких и тяжелых), получила название иммуноглобулинов.
Иммуноглобулины синтезируются в пузырьках пластинчатого аппарата (Гольджи) В-лимфоцитов. По мере созревания пузырек подходит к поверхности клетки, при контакте с ней происходит разрыв наружной мембраны клетки и мембраны пузырька. Свободные иммуноглобулины поступают в плазму, а связанные с мембраной остаются на поверхности клетки. Таким образом, итоговой фазой В-клеточного иммунитета является продукция /g-антител, которая происходит постоянно. Одновременно в организме синтезируются до 105... 107 антител различной специфичности. Интактный В-лимфоцит синтезирует в один час примерно 250...500 молекул IgM и большую часть их отдает в экстрацеллю-лярную жидкость. После специфической антигенной стимуляции и трансформации лимфоцитов в плазматические клетки этот синтез возрастает в тысячи раз.
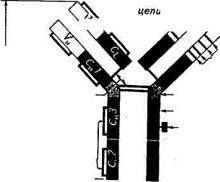
Наиболее полно изучено строение антител, представленных иммуноглобулинами класса G (рис. 4.5).
На схеме видно, что молекула IgG под действием протеолити-ческого фермента папаина расщепляется на три больших фрагмента. Два из них идентичны и обладают свойством взаимодействовать с одной молекулой антигена. Они были обозначены как Fab-фрагменты (Fragment antigen binding), связывающие антиген.
 Гипервариабельные участки легкой цепи Лёгкая м цепь
Гипервариабельные участки легкой цепи Лёгкая м цепь
| (Связывание _ антигена) F°° |
| Гипервариабельные участки тяжелой цепи |
| Межцепьевые " дисульфитные связи |
| Шарнирная область Участок связывания комплемента Углевод |
| (Эффекторные I биологические Fc |
| функции) |
| Внутрицепьевые дисульфидные связи |
| VL и Vfj: вариабельные области Ci и Сн'- константные области |
| Рис. 4.5. Структура молекулы иммуноглобулина G |
Тяжёлая цепь v*v /^Тяжелая цепь
Третий фрагмент, содержащий углеводный компонент, был получен при изучении его свойств в кристаллическом состоянии, за что и назван Fc-фрагментом (Fragment cristalline). В дальнейшем было показано, что Fc-фрагмент взаимодействует с мембранами клеток, выполняя транспортную функцию, фиксацию Cql компонента комплемента и т. д.
Все иммуноглобулины — это тетрамеры, состоящие из четырех полипептидных цепей. Две из них представлены длинными (около 450 аминокислотных остатков), или тяжелыми (heavy), Н-цепями, а две другие — короткими (около 220 аминокислотных остатков), или легкими (light), L-цепями. Легкие цепи являются общими для всех классов и подклассов иммуноглобулинов и могут произвольно сочетаться с любой тяжелой цепью. У большинства позвоночных легкие цепи разделяются по антигенным свойствам на два типа — «каппа» (с-тип) и «лямбда» (1-тип).
Изучение первичной структуры полипептидных цепей иммуноглобулинов показало, что N-концевые участки тяжелых и легких цепей, включающие около 100 аминокислотных остатков, обладают уникальной структурой и аминокислотной последовательностью. Они представляют собой вариабельные области (V-область) тяжелых (VH) и легких (VL) цепей. Указанные вариабельные области, с одной стороны, образуют антигенсвязывающие центры антител, а с другой — определяют их многообразие. Остальная (С-концевая) последовательность тяжелой и легкой цепей идентична у всех молекул данного класса или подкласса, но различна у молекул, относящихся к разным классам и подклассам. Она носит название константная область (С-область). В зависимости от структуры константной области тяжелых цепей молекулы иммуноглобулинов делят на классы. В настоящее время известно пять классов иммуноглобулинов, обозначаемые как IgA, IgM, IgG, IgD и IgE. Тяжелые цепи иммуноглобулинов каждого класса различаются по своим фи-шко-химическим, антигенным и иммунологическим свойствам. Эти цепи обозначаются греческими буквами в соответствии с латинским обозначением класса иммуноглобулинов: IgA (а-цепь), IgM (m-цепь), IgG (g-цепь), IgD (d-цепь) и IgE (е-цепь).
В N-концевой части Н- и L-цепей иммуноглобулинов имеются участки, отличающиеся чрезвычайно высокой вариабельностью (гипервариабельные участки): HI; Н2; НЗ; LI; L2; L3. Каждый из них содержит около шести аминокислотных остатков. Вместе эти участки образуют антигенсвязывающий центр.
Цепи иммуноглобулинов состоят из доменов — структурных единиц, содержащих около 110 остатков аминокислот. Тяжелые цепи образуют четыре домена: один вариабельный и три константных, обозначаемые Сн1; Сц2; СцЗ. Легкие цепи содержат но два домена — вариабельному (VL) и константному (Q,). Каждый домен представляет собой плотно упакованную глобулу, так как аминокислотные последовательности обладают определенным
 сходством друг с другом. И эти участки полипептидной цепи замыкаются внутрицепочной дисульфидной связью.
сходством друг с другом. И эти участки полипептидной цепи замыкаются внутрицепочной дисульфидной связью.
Каждый из Fab-фрагментов состоит из одной L-цепи (VL- и CL-домены) и половины одной Н-цепи (VH- и Сн-Домены), а Fc-фрагмент образован остатками двух Н-цепей, включающих Сн2 и Сн3 домены.
Классы иммуноглобулинов различаются по своим химическим и биологическим свойствам. Так, их различная скорость полимеризации выражается в том, что /gG существуют в виде мономера с молекулярной массой 160 000, тогда как IgA состоит из одной или двух, а IgM, как правило, из пяти соответствующих субъединиц.
Биологические и иммунологические различия между классами иммуноглобулинов имеют большое практическое значение, так как они обусловливают характер реакции, следующей за образованием комплекса антиген — антитело.
Защитное действие антител может выражаться в прямом повреждении микроорганизмов или в нейтрализации синтезируемого ими токсина. Нейтрализация токсина состоит в прямой блокаде токсофорной группы антителами (антигенные и токсинные детерминанты идентичны). Однако связывание токсина и антитела может вызывать и пространственные изменения конфигурации, которые делают невозможной адсорбцию токсина на субстрате — клетке, чувствительной к токсину.
Иммуноглобулины класса G оказывают сильное нейтрализующее действие на токсины и вирусы, а также способны к преципитации, обладают опсонизирующим действием (опсонины — греч. Opsonion — снабжение пищей — антитела, в присутствии комплемента стимулирующие поглощение чужеродных частиц, бактерий, вирусов и их разрушение лейкоцитами), усиливая фагоцитоз. При умеренном нагревании IgG может связываться с антителом, т. е. начинается реакция, названная связыванием комплемента, ответственность за которую несет Fc-фрагмент. При этом кроме непосредственной нейтрализации вирусов и токсинов IgG способствуют также зависящему от комплемента фагоцитозу и внутриклеточной инактивации микроорганизмов. На долю IgG приходится около 80 % всех иммуноглобулинов.
Более сложной, чем при нейтрализации токсинов, представляется роль антител в разрушении и устранении самого возбудителя. У некоторых грамотрицательных бактерий антитела в связи с неспецифически реагирующими факторами комплемента сыворотки могут вызывать бактериолиз. Антитела и комплемент, однако, не вызывают цитолиз непосредственно, а лишь изменяют вначале поверхность бактериальной клетки таким образом, что она становится доступной действию имеющегося в организме фермента лизоцима. Лизоцим реагирует с мурамилпептидом клеточной стенки и вызывает разрыв в одном или нескольких местах поверхности. Повышение давления внутри клетки, дохо-
дящее до 20 ати, приводит к тому, что микроорганизм с поврежденной стенкой лопается.
Другой путь возможного действия антител — опсонизация заключается в изменении поверхности микробной клетки таким образом, что она становится более удобной для поглощения ее фагоцитами. Как и бактериолиз, опсонизация протекает на фоне активации комплемента, вызванной реакцией антиген — антитело. В отличие от цитолиза, требующего для своего осуществления присутствия на бактериальной клетке всех девяти факторов комплемента, в опсонизации участвуют только четыре компонента комплемента Q...C4.
Особенно выражено опсонизирующее действие антител при колиинфекции новорожденных домашних животных. Большое значение имеют опсонизирующие антитела и для защиты против диплококков и стрептококков. Гладкая поверхность капсулы у диплококков мешает их поглощению фагоцитами. Лишь когда под действием антител их поверхность меняется, т. е. становится шероховатой, они могут быть фагоцитированы и затем уничтожены внутри клетки. Стрептококки поддаются фагоцитозу даже при отсутствии специфических антител, но из-за присутствия особого белка, названного М-антигеном, инактивируются не полностью. Часть их снова выходит из фагоцитов и продолжает размножаться дальше. Антитела, направленные против М-антигена патогенного типа стрептококков, способствуют внутриклеточной инактивации этих микроорганизмов.
|
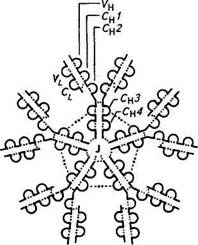
Ведущее место в реакциях бактериолиза и опсонизации принадлежит IgM. Этот класс иммуноглобулинов является самым «древним» в филогенетическом отношении. В ходе иммунного ответа вначале также появляются антитела /gM-клас-са. У новорожденных первые антитела принадлежат к IgM. В сыворотке молекулы IgM существуют в виде пентамера с молекулярной массой 950000. Пять мономерных субъединиц расположены радиально, причем Fc-фрагменты направлены к центру круга, а Fab-фрагменты — кнаружи (рис. 4.6).
Поскольку IgM в полной мере не способны к нейтрализации
токсинов, то животные раннего p„c 4.6. Строение пентамерной структу-
постнатального периода наибо- ры иммуноглобулина м
11 — 3389
лее подвержены токсикоинфекциям. IgM составляют примерно 3...10 % иммуноглобулинов сыворотки крови.
Характерная особенность структуры IgM определяется тем, что она призвана оказывать особое защитное действие против микроорганизмов и других крупных антигенов, имеющих на своей поверхности антигенную мозаику из повторяющихся детерминантов. Антитела IgM связывают соответствующий антиген каждым из своих пяти участков связывания. Такие повторы антигенных детерминантов характерны для О-антигенов из клеточной стенки грамот-рицательных бактерий, жгутиков бактерий и вирусов, имеющих капсиды. Даже если каждый отдельный участок связывания проявляет лишь небольшую авидность, общая авидность всей молекулы повышается из-за суммирования нескольких таких связей.
Если антитела IgM реагируют таким образом с антигеном, то при нахождении антигена близко к поверхности клетки комплемент настолько активируется, что вызывает локализованное повреждение нижележащей клеточной мембраны. Так, единственная молекула IgM в силу активирования комплемента может вызвать лизис клетки (например, бактерии или эритроцита). Антитела класса IgG, напротив, могут вызвать клеточный лизис путем активирования комплемента только в том случае, когда две молекулы антитела вступают в реакцию на поверхности клетки очень близко друг от друга, но для этого необходимо очень много (тысячи) молекул IgG.
Если антиген не связан с поверхностью клетки, а существует в свободном виде с множеством детерминантов, то антитела IgM не имеют никаких преимуществ перед IgG- Напротив, их участки связывания, характеризующиеся низкой авидностью, не могут все связываться с одной и той же молекулой антитела и легко диссоциируют. Поэтому антитела IgG более эффективны при нейтрализации бактериальных токсинов и при защите клеток от адсорбирования на них вируса.
Сравнение активности антител IgG и IgM по отношению к одному и тому же поверхностному антигену сальмонелл показало, что на основе соотношения масс последние в 20 раз активнее при агглютинации [лат. agglutinatio — склеивание в глыбки (комочки) микробов, эритроцитов или других клеток и выпадение их в осадок в присутствии электролитов] сальмонелл, более чем в 100 раз активнее в инактивации микробной клетки с участием комплемента и более чем в 1000 раз эффективнее в опсонизации микроорганизмов и других клеток.
Сывороточные антитела IgA в отличие от IgM и IgG не способны к связыванию комплемента и поэтому не могут вызывать ни иммуноприлипания, ни цитолиза или конглютинации. Считают, что этому классу иммуноглобулинов принадлежит функция защиты организма от проникновения возбудителя и развития инфекции. Этот класс иммуноглобулинов участвует в элиминации пи-
щевых и других чужеродных антигенов, которые могут проникнуть в организм через слизистые оболочки, а также в регуляции иммунного ответа на эти антигены.
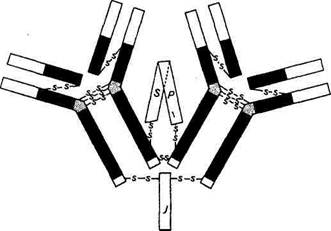
Иммуноглобулины А составляют лишь 10...15 % всех иммуноглобулинов сыворотки. Однако они преобладают в экстра-васкулярных секретах. Большая часть IgA в слюне, слезах, пищеварительных соках, секретах слизистой носа находится в виде секреторного IgA (SIgA), т. е. полимерной формы, состоящей из двух /gA-мономеров, соединяющей молекулы гликопротеина, называемого секреторным компонентом. Биологическая роль секреторного компонента сводится к повышению устойчивости IgA от разрушения протеолитическими ферментами (рис. 4.7).
|
| Рис. 4.7. Схема строения секреторного иммуноглобулина А |
Иммуноглобулин ыЕ — это мономеры, содержание которых в сыворотке крови ничтожно мало — 0,00005...0,0003 г/л, или 0,002 % общего количества Ig. За сутки их синтезируется 0,02 мг/кг; период полураспада в сыворотке крови составляет 2...3 сут, а в коже — 9...14сут. К классу IgE относится основная масса аллергических антител-реагинов. Уровень их значительно повышается у организмов, страдающих аллергией и зараженных гельминтами. IgE связываются с Fc-рецепторами тучных клеток и базофилов. При контакте с аллергеном образуются мостики IgE — антиген — IgE, что сопровождается поступлением ионов кальция в клетку-мишень, активацией в ней биохимических процессов и выделением биологически активных веществ, вызывающих аллергические реакции немедленного типа. Эозинофильный хемотаксический фактор, выделяемый тучными клетками, способствует аккумуляции
эозинофилов и деструкции гельминтов. Предполагается также, что IgE, покрывая паразита, аккумулирует макрофаги благодаря Fc-рецепторам этих клеток.
Иммуноглобулины D — это мономеры; их содержание в крови составляет 0,03...0,04 г/л, или до 1 % общего количества иммуноглобулинов. В сутки их синтезируется от 1 до 5 мг/кг, а период полураспада колеблется в пределах 2...8 сут. IgD участвуют в развитии местного иммунитета, обладают антивирусной активностью, в редких случаях активируют комплемент. Плазматические клетки, секретирующие IgD, локализуются преимущественно в миндалинах и аденоидной ткани. IgD выявляются на В-клетках, отсутствуют на моноцитах, нейтрофилах и Т-лимфоцитах. Полагают, что IgD участвуют в дифференцировке В-клеток, способствуют развитию антиидиотипического ответа, участвуют в аутоиммунных процессах.
Основная масса IgM и IgD находится в плазме, a IgG и IgA распределяются примерно в одинаковых соотношениях между плазмой и межсосудистой тканью.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 970; Нарушение авторских прав?; Мы поможем в написании вашей работы!