КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Особенности размножения птиц
|
|
|
|
Размножение птиц имеет ряд характерных черт, которые прежде всего связаны с условиями развития зародыша. В отличие от млекопитающих животных развитие эмбрионов птиц происходит вне организма матери, причем не в водной среде, как у рыб и амфибий, а на суше. В связи с этим яйца птиц обладают сложной системой яйцевых оболочек, предохраняющих зародыш от повреждений и прежде всего от высыхания. Кроме того, яйца содержат большой запас питательных веществ, необходимых для развития зародыша. В ходе эмбриогенеза значительного развития достигают амнион, аллантоис и желточный мешок, которые берут на себя функции питания зародыша, газообмена и обмена веществ.
Особое значение для ветеринарных специалистов имеет изучение физиологии репродуктивного аппарата домашних птиц, особенно кур-несушек, основная хозяйственно полезная ценность которых связана именно с деятельностью органов размножения.
Половое созревание курицы яйценоских пород завершается к 4...5 мес, более крупных мясных — к 6...8 мес, петушки созревают несколько раньше. Продолжительность половой активности домашней птицы составляет у петуха до 4 лет, у кур до 6, у индюка до 3, у индейки до 5, у селезня и утки до 4, у гусака до 5, у гусыни до 8 лет.
Семенники у самцов птиц расположены в брюшной полости на уровне трех последних ребер. В период половой деятельности семенники становятся крупнее; в извитых канальцах образуются спермин. Спермин через семявыносящие канальцы, которые впа-
дают в придаток семенника, переходят в семяпровод, открывающийся в стенке клоаки сосочком. Придатки семенников развиты слабо, придаточные половые железы у птиц отсутствуют. Совокупительного органа у петуха нет, он вводит семя, прижимая свою клоаку к клоаке самки. У селезней и гусаков копулятивный орган (половой член) состоит из фиброзной ткани, образующей кавернозное тело. На его поверхности спирально проходит семенной желоб, который при эрекции превращается в канал.
Объем эякулята у петуха достигает 0,8 мл, количество сперматозоидов в 1 мл составляет 5...7 млрд, при частом совокуплении число сперматозоидов снижается.
С наступлением линьки половая активность петуха постепенно ослабевает и наступает состояние полового покоя. В отличие от яйцеклеток спермин способны длительное время сохраняться в яйцеводе кур, не теряя своей оплодотворяющей способности. Например, куры несут оплодотворенные яйца еще в течение 20 сут после отсадки самцов, у индеек и гусынь этот срок более продолжителен. У регулярно спаривающихся кур все отделы яйцевода заполнены сперматозоидами. После спаривания спермин не сразу появляются в воронке яйцевода, где происходит оплодотворение. Первые оплодотворенные яйца курица несет лишь на третий день.
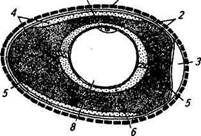
У птиц функционирует только левый яичник; правый, хотя и закладывается, вскоре редуцируется. Соответственно у птиц имеется только один яйцевод, состоящий из воронки, белковой части, перешейка, «матки» («известковой» части) и влагалища, которое открывается в клоаку (рис. 9.9). Общая длина яйцевода у кур-несушек достигает 60 см.
| , поверхностный |

|
Яичник подвешен на брыжейке под передней долей почки. Его глубокий слой образует сосудистую зону слой — фолликулы разной величины и зрелости, прикрепленные на богатых кровеносными сосудами выростах серозной оболочки. В разгар яйцекладки яичник напоминает кисть винограда; у курицы-несушки в нем содержится от 500 до 3500 ооцитов на разных стадиях ооге-неза. Первое мейотическое деление происходит примерно за час до овуляции. Завершение второго мейотического деления и выделение второго направительного тельца происходят уже после разрыва фолликула, попадания яйца в воронку яйцевода и проникновения в яйцо спермия.
Рис. 9.9. Органы размножения курицы:
/ — яичник; 2— воронка яйцевода; 3— белковая часть яйцевода;
4 — перешеек; 5 — «матка»; 6 — прямая кишка (влагалище);
7— клоака
25*
 В ходе своего развития ооциты накапливают в цитоплазме желток, за счет которого достигают значительных размеров (35...40 мм в диаметре у курицы). В созревающих яйцах выделяют первичный и вторичный желток. Первичный желток является производным цитоплазматических структур самого ооцита, а вторичный — результатом секреторной деятельности клеток фолликулярного эпителия. Образование желтка длится около двух недель. Желток куриного яйца состоит из мелких и крупных гранул, более крупные из них называются желточными шарами. Различают белы й и желтый желток, которые различаются по интенсивности окраски и строению желточных шаров. Слои белого и желтого желтков чередуются. В процессе развития ооцита в яичнике курицы слои желтого желтка откладываются в цитоплазме ооцита днем, а слои белого желтка — ночью, когда в яичник поступает меньше питательных веществ из крови.
В ходе своего развития ооциты накапливают в цитоплазме желток, за счет которого достигают значительных размеров (35...40 мм в диаметре у курицы). В созревающих яйцах выделяют первичный и вторичный желток. Первичный желток является производным цитоплазматических структур самого ооцита, а вторичный — результатом секреторной деятельности клеток фолликулярного эпителия. Образование желтка длится около двух недель. Желток куриного яйца состоит из мелких и крупных гранул, более крупные из них называются желточными шарами. Различают белы й и желтый желток, которые различаются по интенсивности окраски и строению желточных шаров. Слои белого и желтого желтков чередуются. В процессе развития ооцита в яичнике курицы слои желтого желтка откладываются в цитоплазме ооцита днем, а слои белого желтка — ночью, когда в яичник поступает меньше питательных веществ из крови.
В яичнике периодически происходит овуляция через полчаса после снесения предыдущего яйца. Длительность этого процесса составляет 1...2 мин. У птиц желтое тело не образуется, поэтому овуляция не задерживается и яйцекладка может быть ежедневной.
После разрыва фолликулярной оболочки ооцит проскальзывает в воронку яйцевода, который при этом перистальтически сокращается, что способствует его продвижению в направлении клоаки. В воронке яйцевода может осуществляться оплодотворение. В этом случае сперматозоиды проникают в яйцо через покрывающие его две нежные пленки. Процесс дробления оплодотворенной яйцеклетки начинается через 4...5 ч после овуляции. Дробление у птиц в отличие от млекопитающих животных частичное дискоидальное. Дробится только зародышевый диск на анимальном полюсе яйца, имеющий ничтожные поверхность и объем по сравнению с недробящейся массой желтка. При попадании яйца в матку дробящийся зародыш состоит из 4...8 бластомеров, а к моменту откладывания их число достигает 256. В таком состоянии яйцо более трех недель может сохранять способность к дальнейшему развитию, если будут созданы соответствующие температурные условия и влажность.
Движению ооцита по яйцеводу способствует мерцательный эпителий стенки яйцевода. Прохождение по верхней части яйцевода до белковой части длится около 15...18 мин. В белковой части, длина которой 30...40 см, формируемое яйцо находится З...3,5ч. При продвижении желток вращается вокруг продольной оси, что происходит благодаря спиральному расположению слизистой оболочки яйцевода. В процессе вращения наслаивается четыре слоя белка, выделяемого железами яйцевода.
Сначала желток покрывается тонким слоем плотного белка, который вследствие вращения желтка образует на его полюсах шнурообразные тяжи — х а л а з ы. Благодаря наличию халаз желток может свободно вращаться таким образом, что при перевора-
чивании яйца зародышевый диск всегда находится сверху, так как ее вегетативная часть тяжелее анимальной (где расположен дробящийся зародыш). Следовательно, при естественном насиживании зародышевый диск всегда смещается ближе к телу наседки.
В белковой части яйцевода наращивается 40...50 % белка, остальной — в перешейке и матке, где яйцо может находиться 1,5...20ч. В перешейке начинается образование подскорлу-повой оболочки, состоящей из двух листков, которые в области тупого конца разделяются, между ними появляется воздушная камера — пуга. После образования подскорлуповой оболочки яйцо принимает форму, характерную для данного вида птицы.
В матке начинается медленное образование на яйце известковой (скорлуповой) оболочки — скорлупы. В период формирования скорлупы усиливается кровоснабжение матки, в результате чего в ее клетки возрастает поступление солей кальция, которые выделяются в полость матки. В скорлупе имеется большое число пор, через которые осуществляются газообмен и испарение воды. Больше пор расположено на тупом конце яйца, в связи с чем при хранении яиц в сухом месте увеличиваются размеры воздушной камеры.
Таким образом, готовое к откладке яйцо окружено целым рядом яйцевых оболочек, через которые осуществляется связь зародыша с окружающей средой — его питание и защита при развитии вне материнского организма (рис. 9.10). Все яйцевые оболочки по происхождению подразделяют на первичные, вторичные и третичные. Первичная оболочка носит название собственно желточной оболочки и является производным цитоплазмы яйцеклетки. Вторичная оболочка представляет собой продукт жизнедеятельности клеток фолликулярного эпителия. Третичные оболочки формируются в результате деятельности желез различных отделов выводящих половых путей — это белковая, подскорлуповая и скор-луповая оболочки.
7 /
|
Рис. 9.10. Схема строения куриного яйца:
/ — скорлупа; 2— подскорлуповые пленки;
I — воздушная камера; 4— белковая оболочка
(белок); 5—халазы; б—желточная оболочка;
7— зародышевый диск; 8— желток
Яйцекладка. Происходит через влагалище — широкое мышечное образование, соответствующее размерам яйца. Во время снесения яйца матка опускается, влагалище и клоака выворачиваются и яйцо выскальзывает наружу через верхний край влагалища, не соприкасаясь ни с влагалищем, ни с клоакой. Этот процесс выведения яйца осуществляется благодаря сокращению мышц клоаки.
Процесс формирования яйца в яйцеводе продолжается от 23 до 30 ч, при этом яйцо более 80 % времени находится в известковой части яйцевода. У большинства кур-несушек овуляция наступает через 10...30 мин после снесения предыдущего яйца. Таким образом, овуляция и оплодотворение происходят с незначительным интервалом, не превышающим 20 мин. Если за этот период оплодотворение не произошло, то яйцо остается неоплод отворенным, даже если в яйцеводе присутствуют сперматозоиды.
Процесс яйцекладки у птиц регулируется нейрогуморальными механизмами. Подобно млекопитающим созревание фолликулов в их яичнике стимулируется гонадотропным гормоном передней доли гипофиза. Еще один гормон гипофиза — лютеинизирую-щий — контролирует процесс овуляции. В самом яичнике также образуются гормоны —это эстрин, или фолликулярный гормон, синтезирующийся в клетках фолликулов, и прогестерон, или гормон желтого тела. Эстрин стимулирует увеличение яйцевода и деятельность его желез. Повышая метаболизм, эстрин регулирует его таким образом, чтобы обеспечить синтез и доставку необходимых для образования яйца веществ.
В естественных условиях яйцекладка у птиц, как правило, бывает весной. Однако у кур благодаря селекционной работе (систематический отбор на яйценоскость) удается вызвать почти непрерывную яйцекладку в течение всего года. Одним из наиболее важных стимулирующих факторов является длина светового дня. Яйцекладка прекращается с наступлением линьки при очень низкой окружающей температуре. Внешние раздражители также оказывают влияние на яйцекладку. Если спугнуть несущуюся курицу, то процесс яйцекладки может значительно задержаться или даже произойти на следующий день, а испуг может не только оттянуть яйцекладку, но и ухудшить качество яйца (наличие кровавых точек). Яйценоскость птиц связана с возрастом производителей: максимальная наблюдается у кур в первый год их жизни, однако яйца мельче стандартов для данной породы. Кроме того, на яйцекладку влияют наследственные признаки данной особи.
Циклом яйцекладки называется период снесения яиц в течение нескольких дней подряд. Циклы могут быть правильными, когда перерыв в яйцекладке происходит через определенное число дней, и неправильными — с перерывами че-
рез разное число дней. У плохих несушек циклы короткие, а перерывы в яйцекладке длинные.
Развитие зародыша у птиц осуществляется благодаря инкубации. Различают естественную инкубацию — развитие зародыша под наседкой (насиживание) и искусственную инкубацию — развитие зародыша в инкубаторе. Для создания оптимального режима в инкубаторах учитывают, что развитие зародышей в яйцах происходит при перемежающейся температуре 37...40°С. При искусственной инкубации продолжительность эмбрионального развития у кур составляет 20...21 сут, у уток и индеек — 27...28, у гусей — 29...30 сут.
Глава 10 ФИЗИОЛОГИЯ ЛАКТАЦИИ
Физиология лактации рассматривает закономерности роста и развития молочной железы, ее взаимодействие с другими системами организма, образование молока и его выведение во время сосания или доения. Лактационная функция присуща только лишь одному, наиболее совершенно организованному классу позвоночных животных — млекопитающим. Способность вскармливать детенышей молоком обеспечивает развивающемуся организму новорожденного наиболее полноценное питание и на самых ранних этапах развития защиту от воздействия неблагоприятных факторов среды.
Строение молочной железы как сложноорганизованного органа, состоящего из ряда тканей (железистой, мышечной, соединительной, жировой и т. д.), обеспечивает оптимальные условия для образования специфического секрета. Железистая, или секреторная, ткань молочной железы у всех видов млекопитающих имеет вид гроздевидной пористой массы, состоящей из ветвящихся протоков, заканчивающихся альвеолярными расширениями. Каждая альвеола представляет собой небольшой пузырек (диаметр от 0,1 до 0,3 мм), стенка которого внутри выстлана одним слоем железистых (секреторных) клеток, а на наружной располагаются мио-эпителиальные клетки, обладающие сократительной способностью. Слепой конец альвеолы имеет округлую форму; на противоположной стороне сужение переходит в выводной проток (рис. 10.1). Молочные альвеолы объединяются в отдельные группы, или дольки, имеющие длину около 1,5 мм, ширину 1 и высоту 0,5 мм; их объем 0,7...0,8 мм3. Тонкие выводные протоки, начинаясь от альвеол, постепенно сливаются с аналогичными протоками соседних альвеол во внутридольковые протоки. Дольки, в свою очередь, объединяются в более крупные образования — доли, имеющие широкие выводные протоки: диаметр долей достигает 2... 17 мм. У коров, коз и овец долевые протоки открываются в своеобразные расширения — молочные цистерны. Крупные выводные протоки существенно отличаются по строению от начальных выводных протоков: слизистая оболочка их выстлана двухслойным цилиндрическим эпителием, а внутренняя стенка имеет складчатую поверхность. За эпителиальным слоем в стенке прото-
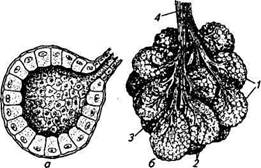
Рис. 10.1. Структура альвеолярного отдела молочной железы:
а —альвеола; б — долька молочной железы; 1 — дольки; 2— внутридольковые протоки; 3— междольковые протоки; 4— выводящие протоки
ка располагаются циркулярно и продольно гладкомышечные волокна. Устья протоков заканчиваются утолщением циркулярного слоя мышц в виде сфинктеров. Молоко, образующееся в альвеолах, по протоковой системе попадает в молочную цистерну — резервуар, который по своему анатомическому строению можно разделить на цистернальный отдел железы и цистерну соска. Наиболее выражена цистерна у коров и коз; у кобыл на один сосок приходится две цистерны. Полость молочной цистерны отделена от цистерны соска круговой сфинктерообразной складкой.
Соединительная ткань, или строма, молочной железы выполняет механическую, поддерживающую и защитную функции. В со-единительнотканой основе проходят кровеносные, лимфатические сосуды, нервы, и она является продолжением соединительно-тканого аппарата брюшной стенки. Брюшная фасция формирует соединительнотканую пластинку, которая, переходя в область основания вымени, образует подвешивающую связку, состоящую из двух листков, и делит вымя на правую и левую половины. Снаружи вымя поддерживается наружной фасцией, представляющей собой продолжение поверхностной брюшной фасции. Соединительная ткань формирует многочисленные перегородки — трабекулы — между долями, дольками и альвеолами. Благодаря оптимальному соотношению эластических и коллагеновых волокон в строме при наполнении вымя больше растягивается в горизонтальном направлении (в стороны), чем в вертикальном (вниз).
Жировая ткань вымени у коров составляет в среднем 43 % органа и развивается независимо от подкожной жировой клетчатки. Она тесно связана с кровеносной системой молочной железы: каждая жировая клетка непосредственно контактирует с петлей
капилляра. Кроме высокой степени васкуляризации в жировой ткани молочной железы мощная сеть симпатической (адренерги-ческой) иннервации способствует быстрой мобилизации жировых запасов, востребуемых при перестройках органа и интенсификации функциональной активности.
Иннервация молочной железы у различных видов животных осуществляется спинномозговыми нервами. У животных, у которых молочная железа представлена компактными образованиями в паховой области (жвачные, непарнокопытные и морская свинка), участвуют в иннервации пояснично-крестцового сплетения: в основном это наружный срамной нерв, в меньшей степени — подвздошно-паховые нервы. У видов животных с множественными железами (большинство грызунов, хищные, свиньи и т. д.) иннервация осуществляется межреберными и поясничными нервами. Молочные железы обладают развитым рецепторным аппаратом, благодаря которому осуществляется тактильная и температурная рецепция; афферентные нервы обеспечивают передачу соответствующей информации в различные отделы ЦНС. Эфферентные нервы (симпатический отдел вегетативной нервной системы) осуществляют как адренергическое, так и холинергическое нервное влияние на течение процессов развития и функционирования молочной железы.
Кровоснабжение молочных желез осуществляется из различных отделов сосудистой системы и имеет видовую специфичность. У коровы, козы и кобылы кровь поступает в основном через наружную срамную артерию, а оттекает по наружной срамной и подкожной брюшной венам. У свиноматки к наружной срамной прибавляются передняя подчревная артерия, которая питает молочные железы, находящиеся краниальнее пупка, и артериальные ветви, доставляют кровь из внутренней и наружной грудных артерий к первым парам грудных желез. Развитие интраорганных артерий тесно связано с развитием железистой паренхимы. У высокопродуктивных коров емкость артериального русла в 4—5 раз выше, чем у малопродуктивных. Артерии I и II порядка имеют обычно бифуркационный тип деления с углом отхождения менее 90°. Артерии сначала следуют по междолевым и междолько-вым пространствам, а затем образуют капиллярную сеть во внут-ридольковой соединительной ткани.
Лимфатическая система молочной железы наряду с венозной является системой оттока и состоит из замкнутых лимфатических капилляров, расположенных в паренхиме, и объединяющих их лимфатических сосудов. Лимфа оттекает по лимфатическим капиллярам и сосудам, проходит через лимфатические узлы и по выносящим лимфатическим сосудам поступает в поясничную лимфатическую цистерну, а из нее по грудному лимфатическому протоку попадает в переднюю полую вену и вместе с кровью в сердце.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 6315; Нарушение авторских прав?; Мы поможем в написании вашей работы!