КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кожный анализатор
|
|
|
|
В г
Рис.15.5. Рефлекс на растяжение мышцы:
А. В исходном «заданном» состоянии груз небольшой массы (1) удерживается экстра-фузальными волокнами мышцы. В нервных волокнах, образующих афферентные окончания, регистрируются только редкие спонтанные потенциалы действия. Б. При увеличении массы груза (2 > 1) мышца с мышечными веретенами растягивается. В афферентных волокнах повышается частота потенциалов действия, которые поступают через синап-тические контакты на а-мотонейроны (показано стрелкой в направлении от мышечного веретена) и возбуждают их. От а-мотонейронов потенциалы действия направляются к экс-трафузальным мышечным волокнам (стрелки в направлении к мышце) и через синаптичес-кие контакты вызывают сокращение мышцы. В. Сокращение мышцы произошло не до заданной длины. Ликвидация «ошибки» осуществляется с помощью фузимоторных гамма-нейронов, образующих двигательные окончания на интрафузальных мышечных волокнах веретен. Г. Мышца возвращается к заданной длине
41—3389
вичные афферентные окончания, и ос-мотонейроны, которые обеспечивают двигательную иннервацию экстрафузальных мышечных волокон. При растяжении мышцы происходит растяжение и мышечных веретен, что сопровождается повышением частоты потенциалов действия в афферентных волокнах. Поскольку афферентные нейроны синаптически связаны в ЦНС с а-мотонейронами, то в последних также повышается частота потенциалов действия. Распространяясь по эфферентным волокнам, потенциалы действия через синаптические окончания вызывают сокращение — укорочение длины мышцы. Уменьшение растягивающего воздействия на интрафузальные волокна снижает частоту потенциалов действия в афферентных нервных волокнах, и система возвращается в состояние, близкое к исходному. Однако полного восстановления первоначальной длины данная система не обеспечивает. Оставшуюся небольшую разницу между исходной длиной мышцы (до растяжения) и длиной после рефлекторного сокращения (ее называют ошибкой) система не может определить. Для этого потребовалось бы звено обратной связи, т. е. мотонейрон с неограниченно высокой чувствительностью. Способствует возврату мышц к исходной «заданной» длине так называемая фузимоторная система, включающая в себя интрафузальные мышечные волокна и фузимоторные (у) мотонейроны, которые образуют на интрафузальных мышечных волокнах двигательные синапсы. Активация этой системы потенциалами действия из двигательных центров анализатора вызывает сокращение концевых участков веретена и тем самым растяжение центрального несокращаюшегося участка, где находятся афферентные первичные окончания. Это приведет к дополнительному увеличению частоты потенциалов действия в афферентном нейроне, которое будет восприниматься а-мотонейро-ном с последующей посылкой к синаптическим окончаниям экстрафузальных волокон эфферентных потенциалов действия. В результате этого в мышце произойдет дополнительное сокращение и исходная длина будет достигнута.
Из вышеизложенного становится ясно, что миотатический рефлекс служит для поддержания постоянной длины мышцы при изменениях воздействующей на нее нагрузки. Этот механизм у животных, как, по-видимому, и у человека, осуществляется без контроля сознания и играет определяющую роль в поддержании позы. Ответственные за положение тела в пространстве мышцы-разгибатели должны иметь определенную заданную длину и в противовес силы тяжести удерживать конечности животного в выпрямленном состоянии.
Сухожильные рецепторы Гольджи соединены с мышечными волокнами последовательно (а не параллельно, как мышечные веретена), поэтому они должны реагировать на изменение напряжения мышцы, а не длины. Было обнаружено, что через тормозные
интернейроны афферентная импульсация i рсигишрмп I <>iii.*i.*m оказывает влияние на а-мотонейроны, снижая vi>">'< m, us,п> ши
ности. Это, например, может проявляться в понижпни... ими
потенциалов действия, посылаемых к синапсам >kci|),h|>v ини.мы* мышечных волокон, что предотвращает чрезмерное пшфнжгнж мышцы. Предполагается также, что сигнализация сухожильными рецепторами о мышечном напряжении а-мотонейронам cnoioit ствует исправлению неточностей в регуляции длины мышц мин татическими рефлексами.
Кожный анализатор имеет самую большую рецептирующую поверхность, которая превышает в несколько тысяч раз, например, площадь сетчатки глаза, обонятельной области и кортиевого органа слухового анализатора. Кожные ощущения у животных весьма многообразны. К ним в первую очередь можно отнести ощущения легкого прикосновения, давления, вибрации, зуда и щекотки, тепла и холода, боли. Действие каждого раздражения, в свою очередь, имеет множество оттенков. Кожа у сельскохозяйственных животных, за исключением небольших участков, покрыта волосами. Она состоит из двух главных слоев: эпидермиса и дермы. В эпидермисе непрерывно происходят деление клеток и миграция их к поверхности. В ходе этой миграции клетки образуют на поверхности слой мертвых уплощенных клеток — роговой слой, который богат кератином и белком, устойчивым к действию воды и различных химических веществ. Под эпидермисом лежит дерма, представляющая собой мягкую и гибкую структуру благодаря толстому слою из коллагеновых и эластических волокон. Многочисленные и разнообразные рецепторные структуры располагаются в обоих слоях кожи. Кроме того, каждый волос кожи имеет свою афферентную иннервацию. Все рецепторы можно разделить на четыре группы. Для первой группы рецепторов — механорецепторы — адекватным раздражителем является энергия механического стимула, механическое смещение кожи или волос; для второй группы — тепловые рецепторы — повышение температуры кожи; для третьей группы— холодовые рецепторы — понижение температуры. Четвертая группа рецепторов—болевые рецепторы — воспринимает весьма сильные механические и температурные раздражители, вызывающие ощущение боли. Все эти рецепторы образованы разветвлениями нервного волокна (дендрита) первого афферентного нейрона. У одних рецепторов разветвленная часть заключена в одну или несколько капсул (инкапсулированные рецепторы), у других терминали лежат свободно в кожных слоях, но они всегда покрыты шваннов-скими клетками.
 15.3.1. МЕХАНОРЕЦЕПТОРЫ КОЖИ
15.3.1. МЕХАНОРЕЦЕПТОРЫ КОЖИ
Все известные механорецепторы по своим функциональным характеристикам можно разделить на несколько групп. В основе классификации механорецепторов лежит их реакция на сту-пенчатообразные механические стимулы, у которых изменяется амплитуда, скорость ее нарастания и спада, длительность и частота Кроме ступенчатообразных стимулов широко применяются синусоидальные механические стимулы. Механорецепторы входящие в состав какой-либо группы, настроены на восприятие определенного параметра механического стимула.

|
В коже обнаружены механорецепторы, по своим функциональным характеристикам сходные с мышечными веретенами, т. е. детекторы амплитуды - скорости (рис. 15.6, А). В покое эти рецептор обладаютСпонтанной активностью. В ответ на постоянное
в
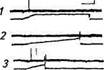
Рис. 15.6. Электрические ответы кожных механорецепторов на механические стимулы различной формы:
А Импульсная активность медленноадаптируюшегося рецептора (дет™ амплитуды-скорости) на ступенчатообразныи механический стимул длительностью около 10 с: /-начало; 2-продолжение, 3- окончание электрического ответа рецептора; калибровка по вертикали 05 мВ, по горизонтали 250 мс. Б. Импульсная активность быст-роадагтшруюшихсяГ механорецепторов (детекторов скорости) на сту-КтообрТзный (I) и пилообразные (2 3, 4) механические стимулы с различной скоростью нарастания; калибровка по вертикали 0,1 мВ, по даонтали для 1 - 125 мс, для 2, 3, 4-250 мс. В Импульсная актив-шсГб^оадаптир^^^^
скоростей) на ступенчатообразный (Г) и пилообразные (2, ^механические стимулы с различной скоростью нарастания; калибровка по вертикали 0,1 мВ, по горизонтали для /, 2— 250 мс, J— n мс
смещение кожи (растяжение или надавлииашн-) они inu-pn руют потенциалы действия, частота которых о. ир.-мпи-м уменьшается (медленная адаптация). При увеличении.imihhiv ды раздражающего стимула частота афферентных иокчппын.ш действия повышается и зависимость между силой рачдрилг ния и частотой импульсации носит логарифмический харак тер Импульсная активность, приходящаяся на передний фрош нарастания раздражения, увеличивается с увеличением скорое ти смещения кожи.
Параллельные морфологические исследования позволили идентифицировать некоторые из этих рецепторов. Ими оказались инкапсулированные механорецепторы - тельца Руффини и тельца Меркеля, а также большая группа свободных нервных окончаний различной степени ветвления (рис. 15.7). Тельца Руффини располагаются в дерме и представляют собой веретенообразные структуры, окруженные капсулой (см. рис. 15.7, Л). Сложнопе-реплетенные коллагеновые волокна являются вспомогательным аппаратом рецептора, они тесно связаны с коллагеновыми пучками лежащими рядом с тельцем. Миелинизированное волокно диаметром 5...10мкм проникает внутрь капсулы, теряет миели-новую оболочку и многократно делится, образуя кустиковидные окончания, располагающиеся на коллагеновых волокнах. У телец Руффини наиболее выраженная электрическая реакция на растяжение кожи. Тельца Меркеля активней реагируют на тактильное (надавливание на кожу) раздражение. Эти структуры встречаются в волосистой коже и представляют собой куполообразные образования диаметром Ю0...500мкм, возвышающиеся над поверхностью кожи (см. рис. 15.7, Б). Рецепторные окончания локализуются непосредственно под базальной мембраной утолщенного эпидермиса. Миелинизированное волокно (средний диаметр 9 5 мкм) также многократно делится и образует дископодобно расширяющиеся окончания с тесно примыкающей к нему клеткой Меркеля. Нервное окончание содержит большое количество
митохондрий. ^^
Кроме медленноадаптирующихся механорецепторов в коже были обнаружены быстроадаптирующиеся рецепторы, которые, как правило, не обладают спонтанной активностью. На постоянное смещение кожи они не генерируют потенциалы действия, какой бы величины оно не было (см. рис. 15.6, Б). Генерация потенциалов действия происходит только во время изменения амплитуды нарастания или спада стимула. С увеличением скорости изменения амплитуды раздражения число потенциалов действия, а следовательно, и их средняя частота увеличиваются. Эти механорецепторы были классифицированы как детекторы скорости. Быстроадаптирующиеся ответы были зарегистрированы от нервных волокон, иннервирующих волосяные луковицы различных типов волос: пуховых, стержневых и вибрисс. Нервные оконча-
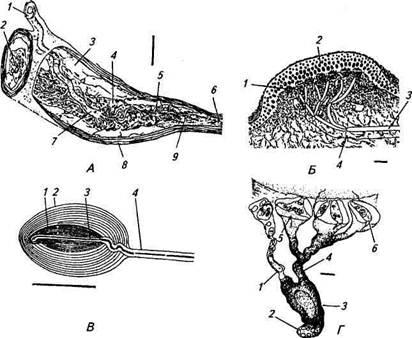
Рис. 15.7. Кожные механорецепторы (графическая реконструкция по серийным срезам):
А. Кожный рецептор растяжения — тельце Руффини: 1 — афферентное нервное волокно; 2, 5— афферентные нервные окончания; 3 — внутрикапсулярное пространство; 4, 6— колла-геновые волокна; 7—место разветвления афферентного нервного волокна; 8— капсула; 9— шванновская клетка с ядром; калибровка 50 мкм. Б. Тактильное тельце — тельце Меркеля: 1 — тактильная клетка (клетка Меркеля) и примыкающее к ней дископодобное нервное окончание; 2 — эпидермис тактильного тельца; 3 — афферентное нервное волокно; 4— немиели-низированное нервное волокно; калибровка 10 мкм. В. Тельце Пачини: 1— внутренняя часть капсулы с наиболее плотным расположением уплощенных слоев клеток (внутренняя колба); 2— наружняя часть капсулы с менее плотным расположением слоев уплощенных эпителиальных клеток; 3 — афферентное нервное окончание; 4— афферентное нервное волокно; калибровка 600 мкм. Г. Свободные немиелинизированные нервные окончания (7 окончаний), образованные разветвлением миелинизированного нервного волокна и располагающиеся около пухового волоса кожи: 1 — одно из афферентных нервных окончаний (продольный срез); 2— семь нервных окончаний, покрытых шванновской клеткой (поперечный срез); 3 — шванновская клетка; 4— четыре нервных окончания, покрытые шванновской клеткой; 5, 6— терминальные части нервных окончаний, окруженные складками из шванновских клеток (поперечный срез): нервные окончания заполнены мелкими митохондриями; калибровка 1 мкм
ния, находящиеся в волосяных фолликулах пуховых и стержневых волос, представляют собой сеть тонких немиелинизиро-ванных окончаний, плотно оплетающих структуры волосяной сумки. Рецепторы волосяной сумки образованы разветвленными (см. рис. 15.7, Г) миелинизированными нервными волокнами диаметром около 6 мкм. Каждый волос иннервируется ответвлениями нескольких волокон. В свою очередь, одно и то же волок-
но может иннервировать много волос. Так, например, в ухе кролика одно волокно может давать афферентные веточки примерно к 300 волосяным фолликулам на площади около 1 см2. Вибриссы являются специальными осязательными волосами и обычно длинней и толще, чем другие волосы. У лошадей и коров вибриссы — это длинные торчащие волосы на морде; у кошек, собак и пушных зверей их называют усами. У лазающих животных они находятся на животе. Корень вибриссы окружен кровеносным синусом. Вибриссы сильнее иннервированы, чем обычные волосы. Свыше 100 нервных волокон могут проникать в сумку вибриссы и образовывать внутри афферентные окончания. Кроме быстроадаптиру-ющихся механорецепторов у вибрисс имеются и медленноадапти-рующиеся механорецепторы. Детекторы скорости были обнаружены в различных участках кожи животных, в том числе в коже репродуктивных органов самцов и сосков молочных желез. Морфологически они представляют собой неинкапсулированные нервные окончания различной формы и сложности ветвления.
Какова же природа адаптации в этих механорецепторах? Надо отметить,что от детекторов скорости не удалось зарегистрировать рецепторный потенциал, поэтому о причине адаптации нельзя сказать что-либо определенное. Некоторые исследователи счита-ют,что адаптация обусловлена особыми механическими свойствами вспомогательного аппарата рецепторов, который не пропускает статическую компоненту механического стимула. Так обстоит дело с быстроадаптирующимся механорецептором — тельцем Пачини. Этот рецептор реагирует на довольно быстрые изменения скорости смещения кожи всего лишь 1...2 потенциалами действия. Данный тип рецепторов получил название детекторов быстрых скоростей, или детекторов толчков.
Тельце Пачини состоит (см. рис. 15.7, В) из нервного окончания, мембрана которого содержит механоактивируемые ионные каналы, и окружающей его капсулы в виде концентрических слоев из уплощенных клеток эпителия, подобных слоям чешуек в луковице. Тельце имеет весьма крупные размеры и может достигать 0,5...0,7 мм в диаметре и 1 мм в длину. Когда тельце при надавливании на него ступенчатообразным механическим стимулом деформируется, эта деформация через капсулу механически передается к мембране нервного окончания. Последняя обычно реагирует кратковременной деполяризацией (рецепторный потенциал), приходящейся на начало и конец раздражения (рис. 15.8, А). Если снять максимально возможное количество оболочек (полностью удалить оболочки капсулы никому не удавалось), то рецепторный потенциал будет приближаться к ступенчатообразной форме (см. рис. 15.8, Б). Была предложена механическая модель капсулы, представляющая ее слои как параллельно и последовательно соединенные вязкоэластичные элементы. Анализ показал, что давление статической (постоянной) части стимула ослабляется

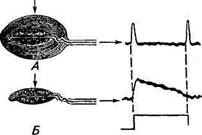 Рис. 15.8. Электрические ответы тактильного тельца Пачини на ступенчатообразное механическое раздражение:
Рис. 15.8. Электрические ответы тактильного тельца Пачини на ступенчатообразное механическое раздражение:
| Стимул |
А. Интактное тельце Пачини. На начало и конец раздражения генерируется кратковременный деполяризационный рецепторный потенциал. Б. У тельца Пачини механически удалены внешние слои капсулы. На механический стимул генерируется более длительный рецепторный потенциал
примерно в 100 раз при достижении области афферентного окончания. Только быстрый компонент, т. е. изменение скорости раздражения, может проходить через капсулу. Таким образом, капсула играет роль механического фильтра. Надо отметить, что в коже коров, коз, лошадей и других животных было обнаружено большое число как телец Пачини, так и других подобных им рецепторов с различными размерами и количествами слоев капсулы.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 1027; Нарушение авторских прав?; Мы поможем в написании вашей работы!