КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Інтеграція живої матерії на різних рівнях організації
|
|
|
|
БІОГЕОЦЕНОЗ ЯК СЕРЕДОВИЩЕ ЕВОЛЮЦІЇ
Згадаєте! Які існують рівні організації живої матерії? Чому популяцію вважають елементарною одиницею еволюції?
Різні рівні організації живої матерії співвідносяться між собою за принципом «мотрійки»: кожний наступний рівень містить у собі всі попередні. Так, з биомолекул складаються клітини, із клітин - організми, з організмів - популяції, а з останніх - біоценози. Вплив середовища на певний рівень здійснюється опосередковано, через усе більше високі рівні. Тому від факторів навколишнього середовища найбільше залежать біоценози, найменш - биомолекулы, що визначає різні ступені інтеграції живих систем кожного рівня.
Інтеграція (від лат. интегратио - поповнення) - структурне об'єднання окремих частин, що входять до складу цілісної системи, і погодженість їхніх дій (напр., клітини в багатоклітинному організмі).
Найвищий ступінь інтеграції здійснюється на молекулярному рівні, де величезна розмаїтість молекул становить кілька типів з'єднань: білки, ліпіди, вуглеводи, нуклеиновые кислоти. Навіть самі незначні зміни структури цих молекул, особливо білків і нуклеиновых кислот, можуть докорінно змінити їхньої властивості. Така інтеграція - необхідна умова для нормального функціонування надзвичайно складних молекулярних систем організмів. На клітинному рівні ступінь інтеграції зменшується: кількість і форма певних органелл може значно варіювати, не порушуючи життєдіяльність клітини.
На организменном рівні інтеграція ще нижче: значно коливаються показники маси, розмірів, фарбування й більшість інших ознак. Ще менш інтегровані популяції, у яких постійно змінюються кількість особин, співвідношення між віковими групами, фазами розвитку й ін. ознаки. Ин-тегрированность у біоценозі дуже слабка й зводиться до підтримки на певному рівні круговороту речовин і перетворень енергії переважно в ланцюгах харчування. Окремі ланки цих ланцюгів можуть досить легко заміщатися іншими. Ступінь інтеграції визначає рівень ефективності використання енергії живою системою. Регуляторні механізми також значно жорсткіше на нижчих рівнях у порівнянні з вищими.
Важливою особливістю живих систем всіх рівнів організацій є їхня здатність нормально функціонувати тільки в стабільних умовах, тому на кожному рівні організації підтримується гомеостаз його внутрішнього середовища. Найбільш нестабільні умови навколишнього середовища й відповідно нестійкий гомеостаз спостерігається в надорганизменных живих систем.
Біогеоценоз як середовище еволюції. Сучасні еволюційні погляди засновані на ідеях В. И. Вернадського (мал. 1) про взаємозв'язок всіх рівнів організації живого в єдиній біосфері.
У кожному біогеоценозі всі популяції взаємозалежні трофічними, просторовими й іншими зв'язками й постійно впливають один на одного. Тому будь-які еволюційні зміни в популяціях викликані факторами, що діють через біогеоценоз як цілісну систему. Отже, біогеоценоз - це середовище еволюційних процесів. Залежно від типу біогеоценозу еволюція вхідних у нього популяцій може відбуватися в різних напрямках.
У стабільні (климаксных) біогеоценозах завдяки великій кількості популяцій, тобто різноманітних екологічних ніш, і розгалуженим трофічним зв'язкам будь-які еволюційні зміни порушують гомеостаз системи, тому елімінація в першу чергу відбувається серед мутантних особин. Такі системи можуть поступово змінюватися як єдине ціле в ході сполученої (когерентної) еволюції. Її прикладом можуть служити пристосування покрытосеменных рослин до запилення комахами, а останніх - до харчування їхнім пилком і нектаром. У біогеоценозах ці відносини ще більш ускладнені: наприклад, комахи-запильники служать їжею для інших тварин; існують комплекси видів, що харчуються квітковими рослинами, при цьому не запилюючи їх. Стабільні біогеоценози становлять основу живого компонента всіх ландшафтно-клима-тических зон.
Виды, з популяцій яких складаються стабільні біогеоценози, називають ценофилами (від греч. койнос — загальний і филео -люблю). Вони здебільшого вузько пристосовані (спеціалізовані) до певних умов того або іншого типу климаксных біогеоценозів і тому екологічно й эволюционно непластичні.
При порушенні стабільного біогеоценозу він відновляється шляхом послідовних змін своїх компонентів - сукцесії. Як вам уже відомо, перші сукцесії збіднені видами, у них відбуваються різкі коливання чисельності, тверда конкуренція й ін. Кожна наступна сукцесія має у своєму складі все більше число видів; так відбувається до повного поновлення видової розмаїтості, тобто відновлення климаксного біогеоценозу. Види, що приймають участь у ранніх сукцесіях, відрізняються високою екологічною пластичністю, мало специализи-
рованы й не зустрічаються в зрілих біогеоценозах. Вони одержали назву ценофобы (від греч. койнос - загальний і фобос - страх).
Звідки ж попадають виды-ценофобы в ділянки зруйнованих біогеоценозів? Існують так звані интразональные біогеоценози, умови існування в які увесь час змінюються, наприклад экосистемы річкових долин. Ріка, вимиваючи породи, постійно поглиблює своє русло; частина вимитих речовин відкладається на її берегах у вигляді піску й глини. У міру поглиблення русла ці наноси покриваються рослинністю, образуя заплавні луги (тобто співтовариства, періодично затоплювані під час паводків), сукупність яких має назву лугова тераса. Коли русло заглиблюється настільки, що частина лугів ніколи не затоплюється, останніх заміщають соснові ліси (т.зв. борові тераси), які, у свою чергу, згодом заміняє листяний ліс (лесові тераси). У таких мінливих умовах біогеоценози перебувають у стані безперервної сукцесії й населені переважно ценофоб-
ными видами.
На границях між стабільними біогеоценозами умови перебування більше різноманітні й тому вони більше багаті видами, чим кожний із сусідніх біогеоценозів. Крім ценофилов з обох стабільних экосистем, отут представлені й ценофобы. Наприклад, біогеоценози лісостепу із цієї причини мають більша видова розмаїтість, чим степової й лісової зон, які вони
розділяють.
Місце виду в організації живої матерії. Як ви помітили,
видовий рівень організації не вписується в систему «мотрійки», тому що різні популяції кожного виду входять до складу різних біогеоценозів. Яка роль видів у біосфері? Міграції особин між популяціями одного виду з різних біогеоценозів змінюють частоти зустрічальності певних станів ознак і генофонди популяцій у цілому. Це відбивається на щільності популяцій і сприяє включенню певних регуляторних механізмів біогеоценозів на її збалансування, тобто веде до ряду мікроеволюційних змін. Отже, популяції одного виду як би «зшивають» окремі біогеоценози в єдину надсистему. Крім того, завдяки міграціям здійснюється горизонтальне переміщення біомаси між біогеоценозами на відміну від вертикального, що відбувається в ланцюгах харчування між популяціями різних видів однієї экосистемы.
На схемі (мал. 131) показаний подібний обмін між різними популяціями трьох видів у дев'ятьох біогеоценозах. Вид 1 здатний до міграцій через долини рік, і тому біогеоценози А и Д, розділені біогеоценозом В, можуть обмінюватися особинами виду 1 через розташовані на іншому березі экосистемы Ж и 3. Види 2 і 3 не можуть переборювати водні перешкоди, тому їхньої популяції на левом і правом берегах ізольований друг від дру-
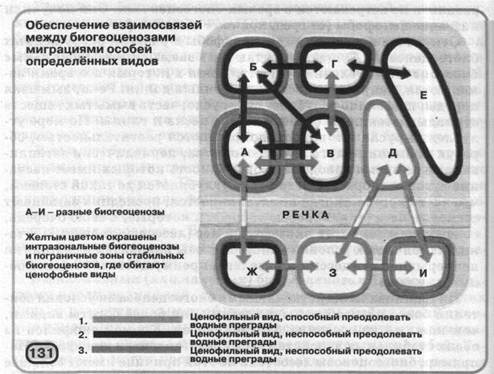
га. На прикладі виду 2 показано, що популяції біогеоценозів А и Г можуть обмінюватися особинами через экосистемы Б и В, а популяції виду 3 біогеоценозів А и Г - тільки через В. Отже, хоча всі три види одночасно присутні тільки в біогеоценозі А, міграції особин кожного з них у сукупності забезпечують зв'язку між всіма дев'ятьма экосистемами.
ВИСНОВКИ
Биоценотический рівень містить у собі послідовно вкладені один в інший рівні організації живої матерії (принцип «мотрійки»): популяционный, организмен-ный, клітинний і молекулярний. Біогеоценоз - це середовище еволюції.
У стабільних біогеоценозах темпи історичних змін невисокі, і популяції різних ценофильных видів еволюціонують сопряженно (когерентна еволюція). У постійно змінюються интразональных біогеоценозах відбувається некогерентна еволюція переважно серед ценофобных видів. Міграції особин між популяціями одного виду змінюють генофонди популяцій, здійснюючи зв'язок різних экосистем.
КОНТРОЛЬНІ ПИТАННЯ:
1. Як змінюються ступені інтеграції й ефективність підтримки гомеостазу від молекулярного до биогеоценотического рівнів організації живої матерії? 2. Що таке ценофобы й ценофилы, когерентна й некогерентна еволюція? 3. Яка роль видів у забезпеченні цілісності біосфери?
Подумайте! Яким образом у сукцесії включаються цено-фобные й ценофильные види?
|
|
|
|
|
Дата добавления: 2014-12-07; Просмотров: 2654; Нарушение авторских прав?; Мы поможем в написании вашей работы!