КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Координационная деятельность центральной нервной системы
|
|
|
|
Классификация нервных центров
Основные функции нервной системы (табл.2.2.)
| Функция цнс | Примеры ее проявления |
| Регуляция деятельности органов и систем | Нервные импульсы, поступающие к сердцу из центра блуждающего нерва, замедляют его работу |
| Координация работы клеток, органов и систем | Во время акта глотания нервная система координирует деятельность языка, надгортанника, мягкого неба и мышц глотки, обеспечивая свободное прохождение пищи из глотки в пищевод. |
| Связь организма с внешней средой | Прикосновение руки к горячему предмету вызывает рефлекторное отдергивание ее от раздражителя |
| Организация высших психических процессов | Нервная система (и особенно ее высший отдел – кора больших полушарий) является материальной основой таких психических явлений, как сознание, мышление и речь. |
2.3.Нервный центр — центральный компонент рефлекторной дуги, где происходит переработка информации, формируются программа действия и эталон результата. Принято различать понятия анатомического и физиологического нервного центра.
Анатомический нервный центр - это совокупность нервных клеток, выполняющих общую для них функцию и лежащих в определенном отделе ЦНС.
В физиологическом отношении нервный центр – это сложное объединение нескольких анатомических нервных центров, расположенных в разных отделах ЦНС и обусловливающих сложнейшие рефлекторные акты.
А.по локализации:
ü Спинальные (расположенные в области спинного мозга)
ü Бульбарные (расположенные в области продолговатого мозга)
ü Мезенцефальные (расположенные в области среднего мозга)
Б.по принадлежности к отделу нервной системы:
ü Соматические
ü Вегетативные
В.по эффектору:
ü Дыхательный
ü Пищеварительный
ü Сосудодвигательный
Поскольку нервный центр состоит из нейронов, синаптически связанных между собой, его основные свойства обусловлены свойствами синапсов.
2.3.2.Свойства нервных центров зависят от их строения и механизмов передачи возбуждения в синапсах. Выделяют следующие свойства нервных центров
1. Одностороннее проведение возбуждения в нервном центре. Возбуждение в ЦНС проводится в одном направлении - с аксона на дендрит или тело клетки следующего нейрона. Одностороннее проведение возбуждения зависит от строения синапса и гуморальной природы передачи в нем импульса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие медиатор, расположены на постсинаптической мембране. Это свойство легко демонстрируется в опыте с раздражением спинномозговых корешков и регистрацией ПД. Если раздражать задние корешки (афферентные пути), то на передних корешках (эфферентные пути) регистрируется ПД. Но при раздражении передних корешков на задних корешках ПД не регистрируется. Следовательно, в ЦНС возбуждение передается только с афферентного пути на эфферентный
2. Замедление проведения возбуждения (центральная задержка). В системе рефлекторной дуги медленнее всего проводится возбуждение в синапсах ЦНС. В связи с этим центральное время рефлекса зависит от количества вставочных нейронов. Чем сложнее рефлекторная реакция, тем больше центральное время рефлекса. Его величина связана со сравнительно медленным проведением возбуждения через последовательно включенные синапсы. Замедление проведения возбуждения создается вследствие относительной длительности осуществляющихся в синапсах процессов: выделение медиатора через пресинаптическую мембрану, его диффузия через синаптическую щель, возбуждение постсииаптической мембраны, возникновение ВПСП и его переход в ПД.
3. Трансформация ритма возбуждения. Нервные центры способны изменять ритм поступающих к ним импульсов(рис.2.2.) Они могут на одиночные раздражители отвечать серией импульсов или на раздражители небольшой частоты - возникновением более частых ПД. В результате ЦНС посылает к рабочему органу количество импульсов, относительно независимое от частоты раздражений. Это связано с тем, что нейрон является изолированной единицей нервной системы, к нему в каждый момент приходит множество раздражений. Под их влиянием происходит изменение мембранного потенциала клетки. Если создается небольшая, но продолжительная деполяризация (длительный ВПСП), то при этом один стимул вызывает серию импульсов.

Рис.2.2. Схема трансформации ритма возбуждения
Причиной трансформации ритма может быть и наличие следового деполяризационного потенциала, который сопровождает ПД и характеризуется определенной степенью деполяризации. На его фоне даже слабые стимулы, приходящие к нейрону, будут вызывать распространяющееся возбуждение. Трансформируется и более частый ритм раздражения. Если к синапсу приходят импульсы с частотой, превышающий его лабильность, то в ответ возникает количество ПД, соответствующее лабильности синапса, т.е. часть импульсов синапс переводит в свой собственный ритм. Это связано с длительностью рефракторного периода и торможением, развивающимся вследствие частого раздражения (часть импульсов попадает в рефракторный период и не вызывает ответа).
4. Ритмическая активность нервных центров (автоматизм). Нервные центры обладают ритмической активностью в разной степени. Различают молчащие нейроны, в которых деполяризация не возникает без действия афферентного стимула, и нейроны, в которых возбуждение возникает без нанесения раздражения. Последние создают фоновую, или спонтанную, активность нервных клеток. Она очень важна, так как эффект раздражения зависит от взаимодействия фоновой активности с пришедшим стимулом. Частота фоновой активности у разных клеток варьирует от нескольких импульсов в секунду до нескольких сотен. Одной из причин фоновой активности является наличие кольцевых связей между нейронами и возможность обратной передачи импульсов по коллатералям одного из нейронов (рис. 3.3.). Ее связывают и со спонтанным выделением «квантов» медиатора нервными окончаниями. Фоновая ритмика имеет большое значение в нормальной деятельности ЦНС, а именно: она расширяет функциональные возможности нейрона. Ритмически активный нейрон имеет повышенную чувствительность к раздражителям. Даже слабое подпороговое раздражение увеличивает его активность, молчащий же нейрон отвечает только на сверхпороговое раздражение. Ритмически активный нейрон отвечает и на стимулирующие, и на тормозящие воздействия, молчащий нейрон не реагирует на тормозное воздействие. Таким образом, фоновая активность придает гибкость и пластичность нервной системе. Высокой ритмической активностью обладают вставочные нейроны.
5. Последействие. Со следовыми явлениями в нервных волокнах связано последействие. Оно проявляется в том, что после прекращения раздражения афферентных нервов по эфферентным путям от ЦНС продолжают следовать импульсы к рабочему органу, вследствие чего рефлекторная реакция некоторое время сохраняется и после выключения раздражения. Последействие зависит от силы раздражения
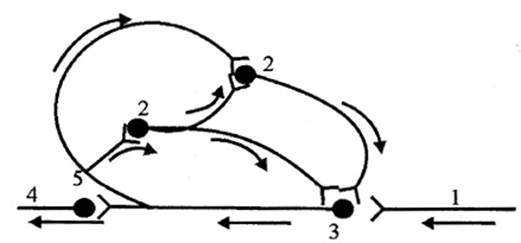
Рис.2.3. Схема кольцевых связей в нервном центре (по Лоренто де Но). Стрелками показано направление движения импульсов: 1 - афферентный путь, 2—промежуточные нейроны, 3—эфферентный нейрон, 4 — эфферентный путь, 5 - возвратная ветвь аксона.
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд могут возникать ПД (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия. Более длительное последействие связано с наличием кольцевых связей между нейронами (рис.2.3.). В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям аксонов к первоначально возбужденному нейрону. От кольцевых связей между нейронами зависит электрическая активность нервных центров, имеющая место и при отсутствии афферентных стимулов, которую называют тонусом центров. Это явление проявляется в том, что нервный центр вследствие спонтанной активности посылает к органам и тканям редкие импульсы и обуславливает их тонус (тонус скелетных мышц, гладких мышц внутренних органов, стенок сосудов и др.). Тоническое состояние центров поддерживается различными гуморальными факторами и афферентными импульсами.
6. Облегчение проведения, или проторение пути. Установлено, что после возбуждения, возникшего в ответ на ритмическое раздражение, следующий стимул вызывает больший эффект или для поддержания прежнего уровня ответной реакции требуется меньшая сила последующего раздражения. Это явление получило название облегчения. Его можно объяснить тем, что при первых стимулах ритмического раздражителя происходит перемещение пузырьков медиатора ближе к пресинаптической мембране и при последующем раздражении медиатор быстрее выделяется в синаптическую щель. Это, в свою очередь, приводит к тому, что вследствие суммации ВПСП быстрее достигается критический уровень деполяризации и возникает распространяющийся потенциал действия.
7. Суммация. Суммация впервые описана И.М. Сеченовым в 1863 г. и заключается в сложении эффектов действия ряда подпороговых раздражителей. Различают два вида суммации: последовательную (временную) и пространственную. Последовательная суммация в синапсах возникает в том случае, когда по одному и тому же афферентному пути к центрам поступает несколько подпороговых импульсов. В результате суммации местного возбуждения, вызванного каждым подпороговым стимулом, возникает ответная реакция. Известно, что один подпороговый афферентный стимул не вызывает ответной рефлекторной реакции, а создает в ЦНС местное возбуждение. При действии серии подпороговых импульсов выделяется достаточное количество медиатора для того, чтобы деполяризация достигла критического уровня и возникло распространяющееся возбуждение. Пространственная суммация заключается в появлении рефлекторной реакции в ответ на два или несколько подпороговых стимулов, приходящих в нервный центр по разным афферентным путям (рис. 2.4.). Пространственную суммацию, как и последовательную, можно объяснить тем, что при подпороговом раздражении, пришедшем по одному афферентному пути, выделяется недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны до критического уровня. Если же импульсы приходят одновременно несколькими афферентным путям к одному и тому же нейрону, в синапсах выделяется достаточное количество медиатора, необходимое для пороговой деполяризации и возникновения ПД.

Рис. 2.4. Схема последовательной (временной) (А) и пространственной суммации (Б)
8. Иррадиация. При возбуждении нейронов нервного центра биоэлектрические импульсы распространяются по коллатералям, вызывая активацию соседних центров. Это явление получило название «иррадиация». В результате при раздражении одного рецепторного поля нередко возникает генерализованная реакция, в которую вовлекаются эффекторы, контролируемые различными нервными центрами (рис.2.5.). Степень иррадиации зависит от количества вставочных нейронов, степени их миелинизации, силы раздражителя.

Рис.2.5. Схема, иллюстрирующая процесс иррадиации возбуждения в нервных центрах: 1, 2, 3 - нервные центры. РО - рабочий орган.
9. Дивергенция и конвергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками в пределах одного или разных нервных центров называется дивергенцией. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих вставочных нейронах. Благодаря этому одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов, что приводит к иррадиации возбуждения. Схождение различных путей проведения нервных импульсов к одному и тому же нейрону получило название конвергенции. Простейшим примером конвергенции является замыкание на одном двигательном нейроне импульсов от нескольких афферентных (чувствительных) нейронов. В ЦНС большинство нейронов получают информацию от разных источников благодаря конвергенции. Это обеспечивает пространственную суммацию импульсов и усиление конечного эффекта (рис.2.6.).

Рис.2.6. Схема, иллюстрирующая дивергенцию и конвергенцию
Явление конвергенции было описано Ч.Шеррингтоном и получило название «воронка Шерринггона», или эффекта общего конечного пути. Данный принцип показывает, как при активации различных нервных структур формируется конечная реакция, что имеет первостепенное значение для анализа рефлекторной деятельности.
10. Окклюзия и облегчение. В зависимости от взаимного расположения ядерных и периферических зон разных нервных центров может проявиться при взаимодействии рефлексов явление окклюзии (закупорки) или облегчения (суммации) (рис.2.7.).

Рис. 2.7. Схема, иллюстрирующая окклюзию и облегчение
При взаимном перекрывании синаптических полей двух нервных центров суммарный результат оказывается меньшим, чем сумма отдельных реакций. Так, при раздражении афферентного поля первого нервного центра возникает условно два двигательных ответа; при активации только второго центра также возникает два двигательных ответа, однако при одновременной стимуляции обоих центров суммарный двигательный ответ равен только трем единицам, а не четырем. Это обусловлено тем, что один и тот же мотонейрон относится одновременно к обоим нервным центрам. В противоположность этому при взаимном перекрывании периферических отделов разных нервных центров мотонейроны, находящиеся в зоне перекрывания и не дающие ответы при изолированном раздражении нервных центров, получают при одновременной стимуляции обоих центров суммарную дозу медиатора, что приводит к пороговому уровню деполяризации. Это вызывает их ответ наряду с мотонейронами, входящими в состав ядерной области, что выражается в усилении суммарного эффекта по сравнению с изолированным раздражением каждого нервного центра в отдельности.
11. Утомляемость нервного центра. Нервный центр обладает малой лабильностью. Он постоянно получает от множества высоколабильных нервных волокон большое количество стимулов, превышающих его лабильность. Поэтому нервный центр работает с максимальной загрузкой и легко «утомляется». Исходя из синаптических механизмов передачи возбуждения, утомление в нервных центрах может объясняться тем, что по мере работы нейрона истощаются запасы медиатора и становится невозможной передача импульсов в синапсах. Кроме того, в процессе деятельности нейрона наступает постепенное снижение чувствительности его рецепторов к медиатору, что называется десенситизацией.
12. Чувствительность нервных центров к недостатку кислорода и некоторым фармакологическим веществам. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходима энергия и постоянный приток нужного количества кислорода. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5-6 минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15-20 минут после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 минут отсутствия кровоснабжения.
13. Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и приспособляемость нервных центров. Это особо ярко проявляется после удаления различных отделов мозга. Нарушенная функция может восстановиться, если были частично удалены какие - то отделы мозжечка или коры больших полушарий. О возможности полной перестройки центров говорят опыты по сшиванию функционально различных центров. Если перерезать двигательный нерв, иннервирующий мышцы конечности, и его периферический конец сшить с центральным концом перерезанного блуждающего нерва, регулирующего внутренние органы, то через некоторое время периферические волокна двигательного нерва перерождаются (вследствие их отделения от тела клетки), а волокна блуждающего нерва прорастают к мышце. Последние образуют в мышце синапсы, свойственные соматическому нерву, что приводит к постепенному восстановлению двигательной функции. В первое время после восстановления иннервации конечности раздражение кожи вызывает свойственную блуждающему нерву реакцию - рвоту, так как возбуждение от кожи по блуждающему нерву поступает в соответствующие центры продолговатого мозга. Через некоторое время раздражение кожи начинает вызывать обычную двигательную реакцию, так как происходит полная перестройка деятельности центра.
2.4.Рефлекторная дуга – путь, по которому проходят нервные импульсы при осуществлении рефлексов. Считается, что у истоков учения о рефлексе стоит Рене Декарт (1596-1650 г). Декарт считал, что в организме есть две разные сущности - тело и душа. Душа является непротяженной сущностью, состоящей из мыслей, разума, ощущений, чувств, то есть всего, что осознается. Отсюда возникло его знаменитое высказывание "Я мыслю, следовательно, я существую"  ("Cogito ergo sum").
("Cogito ergo sum").
Поведение живых существ Декарт рассматривал как механистическое, то есть по аналогии с работой простых механизмов, машин. Он считал, что взаимодействие организма со средой осуществляется при помощи некой "нервной машины", которая состоит из мозга и нервных трубок, расходящихся от него. По нервным трубкам текли "животные духи". Считается, что именно Декарт впервые сформулировал рефлекторный принцип взаимодействия организма со средой, хотя сам термин "рефлекс" появился намного позже. Однако Декарт не распространял рефлекторный принцип на "высший разум", по его мнению, сознательная жизнь людей не подчиняется материалистическим объяснениям. В этом проявляется дуализм Декарта.
Термин "рефлекс" и его биологическое обоснование было предложено в 1800 г чешским физиологом Й. Прохазкой (1749-1820). Именно он развивал представления о рефлексе как механизме приспособления организма к условиям окружающей среды. В отличие от Декарта Прохазка использовал не механистический подход, не идеи и законы механики, а биологический. А также он распространял рефлекторный принцип на все уровни психической деятельности.
Несколько позже, в середине 19 века, были получены новые данные об анатомии спинного мозга. Благодаря этому умозрительные представления о рефлексе превратились в естественнонаучный факт. Английский невролог Ч. Белл (1774-1842) выявил различия в функциях передних и задних корешков спинного мозга. Независимо от Белла к такому же выводу пришел французский физиолог Ф. Мажанди (1783-1855). В результате был сформулирован закон Белла-Мажанди: сигнал входит в спинной мозг по задним корешкам, а выходит из него по передним корешкам. В последующий период на основе работ Ч. Белла, Ф. Мажанди и других, в трудах М. Холла и Й. Мюллера, сосредоточивших свое внимание на изучении структурных, анатомических особенностей нервной системы, складывается анатомический подход к деятельности нервной системы, в это время создается представление об анатомической локализации нервных дуг.
 Из отечественных ученых, внесших огромный вклад в развитие науки о рефлексе, первым был Иван Михайлович Сеченов. И. М. Сеченов распространяет преобразованную — уже не анатомическую, а функциональную — концепцию рефлекса на головной мозг. В 1851 году Сеченов стал студентом медицинского факультета МГУ. Затем он много учился в Германии, работал там с Гельмгольцем (изучал хрусталик глаза). В 1860 году он приезжает в Петербург. Характеристика деятельности головного мозга как рефлекторной означает у И. М. Сеченова прежде всего то, что это деятельность закономерная, детерминированная. Исходной своей естественнонаучной предпосылкой рефлекторная теория И. М. Сеченова имеет положение о единстве организма и среды, об активном взаимодействии организма с внешним миром. Это положение составило первую общебиологическую предпосылку открытия Сеченовым рефлексов головного мозга. Обусловленная внешними воздействиями, рефлекторная деятельность мозга — это тот «механизм», посредством которого осуществляется связь с внешним миром организма, обладающего нервной системой. Свои взгляды Сеченов изложил в книге: «Рефлексы головного мозга», не потерявшей своего значения и сегодня.
Из отечественных ученых, внесших огромный вклад в развитие науки о рефлексе, первым был Иван Михайлович Сеченов. И. М. Сеченов распространяет преобразованную — уже не анатомическую, а функциональную — концепцию рефлекса на головной мозг. В 1851 году Сеченов стал студентом медицинского факультета МГУ. Затем он много учился в Германии, работал там с Гельмгольцем (изучал хрусталик глаза). В 1860 году он приезжает в Петербург. Характеристика деятельности головного мозга как рефлекторной означает у И. М. Сеченова прежде всего то, что это деятельность закономерная, детерминированная. Исходной своей естественнонаучной предпосылкой рефлекторная теория И. М. Сеченова имеет положение о единстве организма и среды, об активном взаимодействии организма с внешним миром. Это положение составило первую общебиологическую предпосылку открытия Сеченовым рефлексов головного мозга. Обусловленная внешними воздействиями, рефлекторная деятельность мозга — это тот «механизм», посредством которого осуществляется связь с внешним миром организма, обладающего нервной системой. Свои взгляды Сеченов изложил в книге: «Рефлексы головного мозга», не потерявшей своего значения и сегодня.
Второй — физиологической — предпосылкой рефлекторной теории явилось открытие Сеченовым центрального торможения. Оно стало первым шагом к открытию внутренних закономерностей деятельности мозга, а открытие этих последних было необходимой предпосылкой для преодоления механистического понимания рефлекторной деятельности по схеме: стимул—реакция, согласно механистической теории причины как внешнего толчка, якобы однозначно определяющего эффект реакции.
Рефлекс головного мозга — это, по Сеченову, рефлекс заученный, т. е. не врожденный, а приобретаемый в ходе индивидуального развития и зависящий от условий, в которых он формируется. Выражая эту же мысль в терминах своего учения о высшей нервной деятельности, И. П. Павлов скажет, что это условный рефлекс, что это временная связь. Рефлекторная деятельность — это деятельность, посредством которой у организма, обладающего нервной системой, реализуется связь его с условиями жизни, все переменные отношения его с внешним миром. собирался пойти по стопам отца. Однако, он, прочитав работу Сеченова "Рефлексы головного мозга", оставил семинарию,
 недоучившись в ней одного года, и решил поступать в
недоучившись в ней одного года, и решил поступать в
Медико-хирургическую академию в Санкт-Петербурге. Первыми
работами Павлова были опыты по исследованию пищеварения. За
открытия механизмов регуляции выделения пищеварительного
сока он в 1904 году был удостоен Нобелевской премии.
Павлов экспериментально подтвердил и дополнил идеи Сеченова.
Он подкрепил идеи Сеченова научной концепцией
условного рефлекса, ввел ее в строгие рамки лабораторного опыта.
Итак, основные принципы рефлекторной теории Павлова-Сеченова следующие:
1. Принцип детерминизма (причинности). Этот принцип означает,
что любая рефлекторная реакция причинно обусловлена, то есть нет
действия без причины. Всякая деятельность организма, каждый акт
нервной деятельности вызван определенным воздействием из внешней
или внутренней среды.
2.Принцип структурности. Согласно этому принципу каждая рефлекторная реакция осуществляется при помощи определенных структур мозга. В мозге нет процессов, которые не имели бы материальной основы. Каждый физиологический акт нервной деятельности приурочен к какой-либо структуре.
3.Принцип анализа и синтеза раздражителей. Нервная система постоянно анализирует (различает) с помощью рецепторов все действующие на организм внешние и внутренние раздражители, и на основе этого анализа формирует целостную ответную реакцию - синтез. В мозге эти процессы анализа и синтеза происходят непрерывно и постоянно. В результате организм извлекает из среды нужную для себя информацию, перерабатывает ее, фиксирует в памяти и формирует ответные действия в соответствии с обстоятельствами и потребностями. Павловская рефлекторная теория преодолела представление, согласно которому рефлекс якобы всецело определяется морфологически фиксированными путями в строении нервной системы, на которые попадает раздражитель. Она показала, что рефлекторная деятельность мозга (всегда включающая как безусловный, так и условный рефлексы) — продукт приуроченной к мозговым структурам динамики нервных процессов, выражающей переменные отношения индивида с внешним миром.

В настоящее время структура рефлекторной дуги дополнена еще одним элементом: обратной афферентацией. Под обратной афферентацией понимаютимпульсацию, поступающую в ЦНС от эффектора и информирующую о степени выполнения полученного приказа.
Для осуществления сложных реакций необходима интеграция работы отдельных нервных центров. Большинство рефлексов являются сложными, последовательно и одновременно совершающимися реакциями. Рефлексы при нормальном состоянии организма строго упорядочены, так как имеются общие механизмы их координации. Возбуждения, возникающие в ЦНС, иррадиируют по ее центрам. Координация обеспечивается избирательным возбуждением одних центров и торможением других.
Координация — это согласованность, слаженность рефлекторной деятельности ЦНС, что обеспечивает реализацию всех функций организма.
Выделяют следующие основные принципы координации:
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 4721; Нарушение авторских прав?; Мы поможем в написании вашей работы!