КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Ионотропные и метаботропные рецепторы. Основные группы синаптически активных веществ (лиганды, агонисты, антагонисты)
|
|
|
|
Общая характеристика и классификация нейромедиаторов. Нейромодуляторы
Для непрерывной передачи импульсов нужно быстро удалять медиатор из белка-рецептора, чтобы следующий квант посредника мог с ним взаимодействовать. Механизмы удаления медиатора из синаптической щели могут быть различные: или путем разрушения медиатора специфическим ферментом, находящимся в синаптической щели, или путем обратного захвата медиатора специальным переносчиком и поступления его назад в пресинаптическое нервное окончание или в глиальную клетку, или же путем разрушения медиатора под действием специфического фермента в синаптической щели, или в постсинаптическом нейроне, или глиальной клетке.
Для выделения везикул с медиатором необходимо определенное время, вследствие чего задержка передачи сигнала составляет 0,2-0,5 мс. Поэтому лабильность синапса составляет не более 100-150 имп/с. При большей частоте импульсов присходит блокировка проведения сигнала. Быстрое утомление синапсов связано с исчерпанием запасов везикул с медиатором в пресинаптическом нервном окончании и необходимости определенного времени для восстановления их запасов.
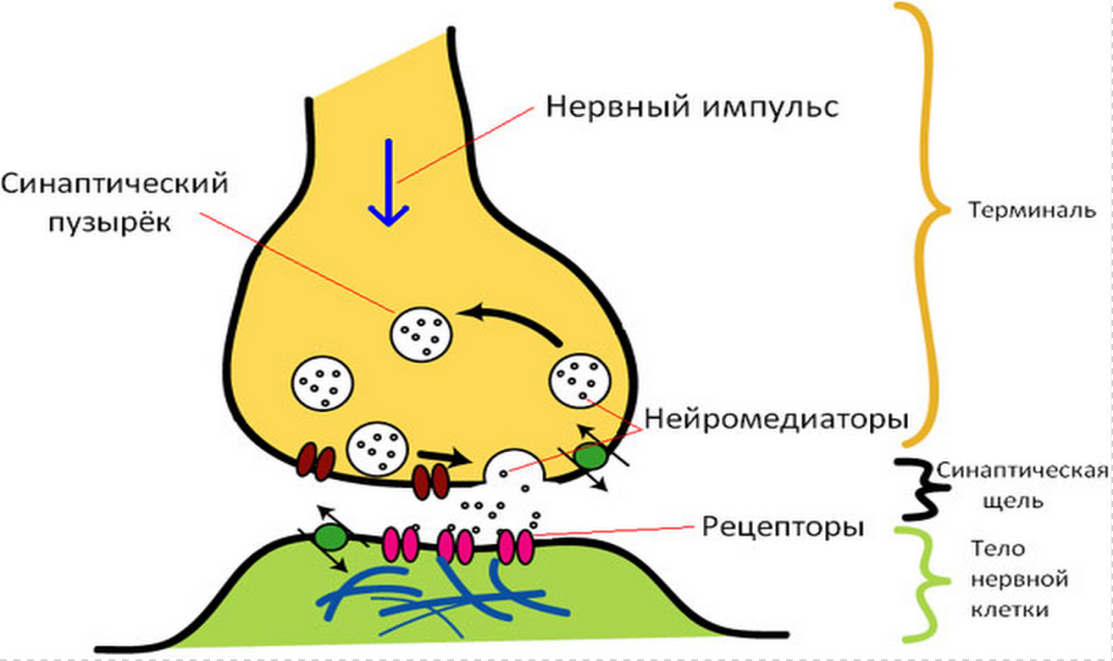
Таким образом, нейромедиаторы – это вещества, образующиеся в пресинаптических нервных окончаниях, хранящиеся там в особых везикулах, выделяющиеся из нервных окончаний под действием нервного импульса в синапс, связывающиеся со специфическим рецептором на постсинаптической мембране и имеющие механизмы для быстрого удаления медиаторов из синаптической щели. В роли медиаторов выступают ацетилхолин, дофамин, норадреналин, серотонин, ГАМК, глицин и некоторые другие вещества.
В зависимости от эффектов на постсинаптической мембране нейромедиаторы делят на возбуждающие и тормозные.
К возбуждающим нейромедиаторам относят ацетилхолин, дофамин, серотонин, глутаминовую кислоту, норадреналин. В некоторых синапсах в качестве возбуждающего нейромедиатора могут выделяться пурины, АТФ, некоторые нейропептиды.
К тормозным нейромедиаторам относятгамма-аминомасляную кислоту (ГАМК) и глицин.
Критерии нейромедиаторов:
1) вещество должно синтезироваться и накапливаться в пресинаптических нервных окончаниях, из которых должно выделяться в ответ на деполяризацию;
2) выделившийся нейромедиатор должен оказывать постсинаптическое действие путем взаимодействия со специфическим постсинаптическим рецептором;
3) вещество должно или разрушаться в синаптической щели, или удаляться из нее с помощью механизма обратного захвата;
4) введение этого вещества в нервную ткань должно оказывать такое же действие, как и эндогенное вещество.
Кроме медиаторов, в синапсе могут выделяться также нейромодуляторы. Нейромодуляторы – это вещества, которую прямо не изменяют состояние синаптических мембран, но влияют на интенсивность и продолжительность действия классических нейромедиаторов и таким образом влияют на активность синаптической передачи сигнала. Чаще всего в роли нейромодуляторов выступают нейропептиды. Нейропептиды – это обширная группа короткоцепочечных пептидов, которые могут выступать и как нейромедиаторы, и как нейромодуляторы.
Например, энкефалины и эндорфины связываются со специфическими рецепторами (с которыми также взаимодействует морфин), при этом происходит подавление чувства боли. Другой пептид – вещество Р, подавляет ощущение боли, а также вызывает сокращение гладких мышц. Ангиотензин II – гормон местного действия, сильно влияет на кровеносные сосуды и работу ЦНС. Так же действует и вазоактивный кишечный пептид (ВИП).
Простагландины действуют как локальные химические агенты, так как они очень быстро инактивируются. Модулируя синаптическую передачу, они влияют на многие физиологические процессы, например, меняют секрецию медиаторов, работу аденилатциклаз.
Рецепторы к нейромедиаторам располагаются на мембранах нейронов или клеток-мишеней (мышечные или железистые клетки). Их локализация может быть и на постсинаптических, и на пресинаптических мембранах. На пресинаптических мембранах чаще располагаются так называемые ауторецепторы, которые регулируют выделение этого же медиатора из пресинаптического окончания. Но есть и гетероауторецепторы, которые также регулируют выделение медиатора, но в этих рецепторах выделение одного медиатора регулирует другой медиатор или нейромодулятор.
Большинство рецепторов – это мембраносвязанные олигомерные белки, которые связывают лиганд (нейромедиатор) с высоким сродством и высокой селективностью. В результате этого взаимодействия запускается каскад внутриклеточных изменений. Рецепторы характеризуются сродством к лиганду, количеством, насыщаемостью и способностью к диссоциации рецептор-лигандного комплекса. У некоторых рецепторов обнаружены изоформы, которые различаются сродством к определенным лигандам. Эти изоформы могут находиться в одной и той же ткани.
Лиганды - это вещества, избирательно взаимодействующие с данным рецептором. Если фармакологическое вещество активирует данный рецептор, оно является агонистом для него, а если снижает его активность – то антагонистом.
Связывание лиганда с рецептором приводит к изменению конформации рецептора, вседствие чего или открываются ионные каналы, или запускается каскад реакций, приводящих к изменениям метаболизма.
Выделяют ионотропные и метаботропные рецепторы.
Ионотропные рецепторы. Вследствие образования постсинаптического потенциала происходит открытие соответствующего ионного канала или сразу при действии медиатора, или через активацию G-белка. При этом рецептор или сам образует ионный канал, или связан с ним. После присоединения лиганда и активации рецептора происходит открытие канала для соответствующего иона. В результате на мембране образуется постсинаптический потенциал. Ионотропные рецепторы – это путь быстрой передачи сигнала и образования ПСП без изменения процессов метаболизма в клетке.
Метаботропные рецепторы. Это более сложный путь передачи сигнала. При этом после связывания лиганда с рецептором происходит активация каскада фосфорилирование-дефосфорилирование. Это осуществляется или прямо, или через вторичные посредники, например, через тирозинкиназу, или через цАМФ, или цГМФ, или инозитолтрифосфат, или диацилглицерол, или за счет увеличения внутриклеточного кальция, что в результате приводит к активации протеинкиназ. Фосфорилирование чаще всего включает в себя активацию цАМФ-зависимой или диацилглицерол-зависимой протеинкиназы. Эти эффекты развиваются более медленно и длятся более долго.
Сродство рецептора к соответствующему нейромедиатору может меняться так же, как и к гормонам, например, за счет аллостерических изменений рецептора или других механизмов. Поэтому сейчас рецепторы обозначают как мобильные и легко изменяемые структуры. Входя в состав мембраны, белки-рецепторы могут взаимодействовать с другими мембранными белками (так называемая интернализация рецепторов). Нейромодуляторы, как и нейромедиаторы, могут влиять на число и чувствительность рецепторов. Длительное присутствие больших количеств нейромедиатора или нейромодулятора может снижать их чувствительность (даун-регуляция), а недостаток лигандов повышать их чувствительность (ап-регуляция).
|
|
|
|
|
Дата добавления: 2014-12-23; Просмотров: 17315; Нарушение авторских прав?; Мы поможем в написании вашей работы!