КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Конкуренция
|
|
|
|
Классификация взаимоотношений
Темы докладов на семинарских занятиях
1. Различия динамики популяций животных и растений.
2. Факторы, влияющие на демографические параметры населения страны.
3. Успехи и проблемы изучения возрастного состава популяций растений.
Глава 8. Взаимоотношения популяций
Взаимодействие популяций (и видов, которые они представляют) – это вопрос, который с равным успехом может рассматриваться в рамках популяционной экологии и при изучении экосистем. С одной стороны, взаимоотношения являются биотическим фактором, влияющим на организм (это и есть влияние «друзей» и «врагов», по определению Э.Геккеля). С другой стороны, взаимодействия организмов – это ячейки системы связей, формирующих экосистему, определяющих распределение ресурсов между организмами одного трофического уровня и закономерности передачи вещества и энергии с одного трофического уровня на другой.
Взаимоотношения организмов разнообразны (табл. 8). Они разделяются на горизонтальные – между организмами одного трофического уровня (как внутри вида, так и между видами) и ветрикальные – между организмами разных трофических уровней. Взаимоотношения первого рода, как правило, носят характер конкуренции, но могут на некоторых этапах жизни организмов быть мутуализмом (т.е. взаимопомощью). Взаимоотношения второго рода более разнообразны: «растение – фитофаг», «хищник – жертва» (иногда эти два типа взаимоотношений объединяют, так как растения по существу тоже жертвы), «паразит – хозяин», мутуализм, комменсализм, аменсализм.
Таблица 8 Типы взаимоотношений видов в экосистеме
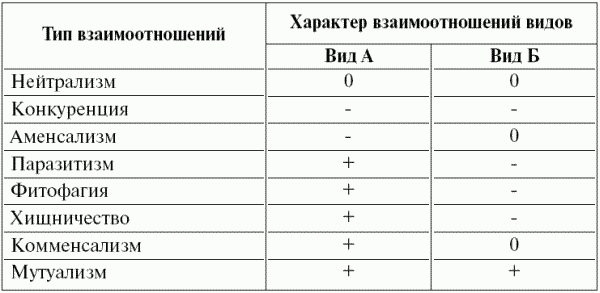
Примечание. Использованы следующие обозначения: 0 – отсутствие взаимоотношений, плюс – положительное влияние, минус – отрицательное влияние.
Кроме материальных взаимоотношений (конкуренции за ресурсы или передачи вещества и энергии при хищничестве или паразитизме), возможны сигнальные (информационные) взаимоотношения. Однако эти сигнальные взаимоотношения тесно переплетены с материальными и лишь корректируют распределение ресурсов между особями или передачу вещества и энергии с одного трофического уровня на другой.
Разделение взаимоотношений в естественных экосистемах по «полезности» и «вредности» некорректно: любые взаимоотношения помогают поддерживать экологическое равновесие и в конечном итоге являются «полезными» для всех видов, которые входят в состав экосистемы. О полезности и вредности отношений между организмами можно говорить только в том случае, если в эти отношения вмешивается человек.
Понятие «экологическое равновесие» подвергается жесткой критике некоторых экологов, которые считают, что этот феномен не существует в природе, а само понятие уходит корнями в представления Карла Линнея о божественной гармонии природы (см. 1.1). Оппоненты понятия «экологическое равновесие» считают, что любые взаимоотношения организмов и любая экосистема как сумма этих взаимоотношений всегда динамичны, так как испытывают влияние множества факторов.
Однако мы полагаем, что понятие экологического равновесия, если оно не абсолютизируется, а рассматривается как некоторая условность (подобная «идеальному газу» или «ускорению» под действием силы земного притяжения), полезно. Системы «хищник – жертва», «паразит – хозяин» и более сложные элементы экосистемы, такие, как пищевые цепи, в состоянии экологического равновесия характеризуются соотношением численности и биомассы взаимодействующих организмов, которые колеблются вокруг некоторой средней величины, соответствующей «экологическому равновесию». Это понятие необходимо при разработке системы рационального использования и охраны природы. Возможно, что более точно этот феномен отражают термины «динамическое равновесие» или «подвижное равновесие».
Контрольные вопросы
1. Назовите основные типы вертикальных и горизонтальных отношений между видами.
2. Почему некорректно разделять взаимоотношения популяций в природе на «полезные» и «вредные»?
3. Что понимается под экологическим равновесием?
Конкуренция – это соревнование организмов одного трофического уровня (между растениями, между фитофагами, между хищниками и т.д.) за потребление ресурса, имеющегося в ограниченном количестве. Д.Тилман (Tilman, 1982, 1983) подчеркивает, что особую роль играет конкуренция за потребление ресурсов в критические периоды их дефицита (например между растениями за воду в период засухи или хищниками за жертвы в неблагоприятный год). Конкуренция играет большую роль в определении видового состава экосистем.
Принципиальных различий у межвидовой и внутривидовой (внутрипопуляционной) конкуренции нет. Возможны как случаи, когда внутривидовая конкуренция является более острой, чем межвидовая, так и наоборот. При этом интенсивность конкуренции внутри популяции и между популяциями может меняться в различных условиях. Если условия неблагоприятны для одного из видов, то конкуренция между его особями может усиливаться. В этом случае он может быть вытеснен (или чаще – потеснен) видом, для которого эти условия оказались более подходящими.
Однако в многовидовых сообществах пар «дуэлянтов» чаще всего не образуется, и конкуренция носит характер диффузной: много видов одновременно конкурируют за один или несколько факторов среды. «Дуэлянтами» могут быть лишь массовые виды растений, которые делят один и тот же ресурс (например, деревья – липа и дуб, сосна и ель и т.д.).
У растений возможна конкуренция за свет, за ресурсы почвы и за опылителей. На почвах, богатых ресурсами минерального питания и влагой, формируются густые сомкнутые растительные сообщества, где лимитирующим фактором, за который конкурируют растения, является свет. При дефиците в почве влаги или элементов минерального питания полог растений бывает разомкнутым и они не конкурируют за свет, а соревнуются за потребление почвенных ресурсов (рис. 19).

Рис. 19. Основной комплексный градиент конкуренции растений (по Tilman, 1988).
При конкуренции за опылителей побеждает тот вид, который более привлекателен для насекомого. Так одним из факторов быстрого распространения в Европе гималайского заносного вида недотрога железконосная (Impatiens glandulifera) является то, что он продуцирует больше нектара, чем его конкуренты в тех же влажных местообитаниях – чистец болотный, дербенник иволистный. Кроме того, нектар недотроги слаще (Chittka, Schurkens, 2001).
У животных конкуренция происходит за ресурсы пищи, например травоядные конкурируют за фитомассу При этом конкурентами крупных копытных могут быть насекомые, подобные саранче, или мышевидные грызуны, способные в годы массового размножения уничтожить большую часть травостоя. Хищники конкурируют за жертвы.
Поскольку количество пищи зависит не только от экологических условий, но и от площади, где воспроизводится ресурс, конкуренция за пищу может перерастать в конкуренцию за занимаемое пространство, т.е. быть не только эксплуатационной, но и интерференционной (см. 8.2). Снижение конкуренции в этом случае возможно при разделении территории на «охотничьи наделы» или на «загоны для выпаса». К примеру, косяки – семейные группы башкирской лошади, в поведении которых сохранились черты их диких предков, рассредоточиваются по «загонам для выпаса», что снижает конкуренцию за фитомассу и способствует равномерному использованию травостоев.
По этой причине лошади – это идеальные фитофаги для особо охраняемых территорий в степной зоне.
Большую роль в разделе территории играют сигнальные взаимоотношения (см. 8.8).
Как и в отношениях между особями одной популяции, конкуренция между видами (их популяциями) может быть симметричной или асимметричной. При этом ситуация, когда условия среды одинаково благоприятны для конкурирующих видов, встречается довольно редко, и потому отношения асимметричной конкуренции возникают чаще, чем симметричной.
При флюктуирующих ресурсах, что обычно в природе (увлажнение или элементы минерального питания для растений, первичная биологическая продукция для разных видов фитофагов, плотность популяций жертв для хищников), поочередно получают преимущества разные конкурирующие виды. Это также ведет не к конкурентному исключению более слабого, а к сосуществованию видов, которые поочередно попадают в более выгодную и менее выгодную ситуацию. При этом ухудшение условий среды виды могут переживать при снижении уровня метаболизма или даже перехода в состояние покоя.
Кроме того, на исход конкуренции часто влияет то, какой из видов первым начал заселять экотоп (принцип лотереи, см. 12.8). Это особенно характерно для маловидовых сообществ водных растений, где более слабый вид может удерживать занятое место, если ему повезло и он его занял первым. Впрочем, лотерея может влиять и на состав сообществ с большим числом претендентов на свободное место. В тропическом лесу на одном гектаре может быть до 150 видов деревьев, и потому занять место выпавшей особи могут представители разных видов.
Влияет на исход конкуренции и то, что победить в конкурентной борьбе больше шансов имеет популяция, в составе которой больше особей и которая, соответственно, будет более активно воспроизводить «свою армию» (так называемый масс‑эффект).
Наконец, конкуренция между видами протекает на фоне отношений с организмами других трофических уровней (хищниками и паразитами). Это также влияет на исход конкуренции, так как более привлекательный как пищевой ресурс вид имеет меньше шансов победить в конкуренции. В итоге в естественных экосистемах виды сосуществуют даже при наличии асимметричной конкуренции, которая должна была бы привести к вытеснению одного из видов. Конкурентное исключение чаще всего наблюдается только в искусственных условиях «микрокосма», когда два конкурирующих вида изолированы и помещены в условия стабильной среды (например в смешанном посеве двух культурных растений с разными конкурентными возможностями).
В естественных экосистемах существуют и специальные механизмы, которые снижают конкуренцию. Главный механизм – дифференциация экологических ниш (см. 9), при которой разные организмы используют разные ресурсы.
Контрольные вопросы
1. Дайте определение конкуренции.
2. Имеются ли принципиальные отличия внутривидовой конкуренции от межвидовой?
3. Какая конкуренция называется асимметричной?
4. Что такое диффузная конкуренция?
5. Приведите примеры конкуренции животных за разные ресурсы?
6. За какие ресурсы среды конкурируют растения?
7. Чем отличаются эксплуатационная и интерференционная виды конкуренции?
8. Как влияет на конкуренцию «принцип лотереи»?
9. Как влияет на конкуренцию «масс‑эффект»?
10. Какое значение для конкуренции имеют флюктуации количества ресурсов?
8.3. Взаимоотношения «фитофаг – растение»
Взаимоотношения «фитофаг – растение» – это первое звено пищевой цепи, в котором вещество и энергия, накопленные продуцентами, передаются консументам.
Для растений в равной мере «невыгодно», чтобы их съели до конца или не съели вовсе. По этой причине в естественных экосистемах проявляется тенденция формирования экологического равновесия между растениями и поедающими их фитофагами. Для этого растения:
– защищаются от фитофагов колючками, образуют розеточные формы с прижатыми к земле листьями, малодоступными для пасущихся животных;
– защищаются от полного выедания биохимическим путем, продуцируя при усилении поедания токсичные вещества, которые делают их менее привлекательными для фитофагов (это особенно характерно для медленно растущих патиентов). У многих видов при их поедании образование «невкусных» веществ усиливается;
– выделяют запахи, отпугивающие фитофагов.
Защита от фитофагов требует значительных затрат энергии, и потому во взаимоотношениях «фитофаг – растение» прослеживается трейдофф: чем растение быстрее растет (и соответственно, чем лучше условия для его роста), тем оно лучше поедается, и наоборот, чем растение медленнее растет, тем оно менее привлекательно для фитофагов. Интенсивное отрастание позволяет растениям с высокой поедаемостью сохраняться и даже доминировать в сообществах.
В то же время эти средства защиты не обеспечивают полную сохранность растений от фитофагов, так как это повлекло бы за собой ряд нежелательных последствий для самих растений:
– несъеденная степная трава превращается в ветошь – войлок, который ухудшает условия жизни растений. Появление обильного войлока ведет к накоплению снега, задержке начала развития растений весной и как итог – к разрушению степной экосистемы. Вместо степных растений (ковылей, типчака) обильно развиваются луговые виды и кустарники. У северной границы степи после этой луговой стадии вообще может восстановиться лес;
– «лишние» листья многих видов трав и кустарников делают крону чрезмерно густой, что ухудшает условия для фотосинтеза (затененные листья «паразитируют», т.е. тратят на дыхание больше органического вещества, чем производят его в процессе фотосинтеза). Этот феномен, который исследовал А.А. Любищев, объясняет благоприятное влияние на урожайность посевов присутствия некоторого количества «вредителей»‑фитофагов, которые осветляют полог растений. Поедание некоторого количества листьев побегов как бы запрограммировано у культурных растений в «память» о своем диком прошлом;
– в саванне уменьшение потребления побегов деревьев веткоядными животными (антилопами, жирафами и др.) приводит к тому, что их кроны смыкаются. В итоге учащаются пожары и деревья не успевают восстанавливаться, саванна перерождается в заросли кустарников.
Кроме того, при недостаточном потреблении растений фитофагами не освобождается место для поселения новых поколений растений.
Подобным образом регулируется равновесие между популяциями видов фитопланктона и зоопланктона. Активно поедаемые водоросли быстро размножаются. Некоторые водоросли, напротив, защищаются от выедания специальными выростами на твердых панцирях (как диатомовые) или объединением в большие колонии, которые не могут быть отфильтрованы рачками. Колониальными формами представлено большинство видов цианобактерий. Защите водорослей от выедания помогает их способность образовывать покоящиеся стадии, играющие ту же роль, что и банки семян у растений. Наконец, некоторые водоросли заглатываются планктонными животными‑фитофагами, но не перевариваются и выделяются с экскрементами живыми.
«Несовершенство» отношений «фитофаг – растение» приводит к тому, что достаточно часто случаются кратковременные вспышки плотности популяций фитофагов и временное угнетение популяций растений, вслед за которыми следует и снижение плотности популяций фитофагов. Например в степях Монголии один раз в 5‑7 лет наблюдаются вспышки численности популяций полевки Брандта, которая полностью выедает надземные части степных растений и разрывает дернину. В итоге угнетаются популяции ковылей, а корневищные злаки быстро разрастаются за счет банка вегетативных зачатков (почек на корневищах). Вслед за вспышкой численности наступает депрессия популяций грызунов из‑за массового заболевания, а популяции ковылей восстанавливаются.
Для обеспечения экологического равновесия в паре «фитофаг – растение» адаптаций растений и животных, как правило, недостаточно. Оно возможно только в случае, если имеется зоофаг, который контролирует плотность фитофага.
По этой причине невозможно экологическое равновесие, если из экосистемы исчез хищник или в нее внедрился вид‑фитофаг, для которого нет контролирующего его хищника.
При разведении сельскохозяйственных животных плотность популяций фитофагов регулируется человеком, который сам устанавливает норму потребления фитомассы животными. В большинстве случаев при этом норма оказывается завышенной, что ведет к развитию процессов пастбищной дегрессии в экосистемах (см. 12.7).
Контрольные вопросы
1. Расскажите об адаптациях, которые позволяют растениям защищаться от фитофагов?
2. Как фитофаги преодолевают «оборону» растений?
3. Почему для устойчивого экологического равновесия в звене пищевой цепи «растение – фитофаг» необходим хищник или паразит?
8.4. Взаимоотношения «хищник – жертва»
Взаимоотношения «хищник – жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка.
Как и при отношениях «растения – фитофаг», ситуации, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами, исключающими полное истребление жертв. Так жертвы могут:
– убегать от хищника. В этом случае в результате адаптации повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей («принцип Тома и Джерри»);
– приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий (например красный цвет, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в листве, а зимой – на фоне белого снега;
– распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
– прятаться в укрытия;
– переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.).
В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки).
В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.).
Полной защиты от хищников всех особей в популяциях животных‑жертв также не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге – к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения их численности подрывается кормовая база.
И.В. Стебаев и др. (1993) образно сравнивают отношения видов‑хищников и их жертв с отношениями между двумя флотами, каждый из которых постоянно усиливает свою огневую мощь и наращивает толщину брони. Но эти флоты никогда не вступают в генеральное сражение.
Тем не менее даже при столь богатом арсенале средств защиты у жертв и нападения у хищников экологическое равновесие в паре «хищник – жертва» только за счет их биотических потенциалов невозможно. И в этом случае, как и в отношениях фитофагов и растений, необходимо третье звено пищевой цепи – естественный «враг» хищника – хищник более высокого порядка или паразит.
В зависимости от характера жертвы и типа хищника (истинный, убивающий жертву сразу, или пастбищник, использующий ее многократно, как, например, оводы или слепни) возможна разное соотношение динамик их популяций. При этом картина осложняется тем, что хищники редко бывают монофагами (т.е. питающимися одним видом жертвы по типу «преследования»). Чаще всего, когда истощается популяция одного вида жертвы и ее добыча требует слишком больших затрат сил, хищники переключаются на другие виды жертв. Кроме того, одну популяцию жертв может эксплуатировать несколько видов хищников и паразитов.
По этой причине часто описываемый в экологической литературе эффект зависимости численности популяций жертв и хищников – пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки – Вольтерры»), – выражен далеко не во всех парах «хищник – жертва». П.В. Турчин (2002) считает осцилляторный (колебательный) характер динамики системы «ресурс – потребитель», которая включает отношения типа «фитофаг – растение» и «хищник – жертва», третьим законом популяционной экологии.
При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага. При этом он должен руководствоваться экологическими нормативами «макисмально допустимого урожая, МДУ», отражающими скорость восстановления плотности популяций. К примеру, без риска разрушить популяцию северного оленя из нее ежегодно можно изымать до 40% животных, а из популяции лосей – только 20%. Примерно также показатель МДУ используется при эксплуатации популяций растений.
Контрольные вопросы
1. Как жертвы «обороняются» от хищников?
2. Как хищники совершенствуют систему преследования жертв?
3. Какие дополнительные условия необходимы для формирования экологического равновесия в паре «жертва – хищник».
4. Почему «эффект Лотки–Вольтерры» в природе проявляется не всегда?
5. Что такое МДУ?
8.5. Взаимоотношения «паразит – хозяин»
Паразиты – это организмы, которые питаются за счет организма‑хозяина. Это очень разнообразная группа организмов (животные, растения, грибы, бактерии), которую изучает специальная наука – паразитология.
Паразиты не убивают хозяина, а длительное время используют его как пищевой ресурс и убежище, лишь сокращая длительность жизни и плодовитость «организма‑дома». Паразиты близки к хищникам‑пастбищникам, но отличаются тем, что если хищник‑пастбищник использует несколько жертв, то паразит связан с одним организмом‑хозяином (а если с несколькими, то меняет их в ходе жизненного цикла).
Различаются следующие группы паразитов.
Биотрофы и некротрофы. Биотрофы всю жизнь питаются за счет живого хозяина, а некротрофы губят его (или часть его тела, например лист растения) и завершают свою биографию как рассматриваемые далее детритофаги.
Микропаразиты и макропаразиты. Различаются по размерам. К микропаразитам относятся вирусы, бактерии, микроскопические грибы и простейшие, к макропаразитам – все прочие.
Истинные паразиты и паразитоиды. Истинные паразиты всю свою жизнь питаются за счет организма‑хозяина (или нескольких хозяев, если в течение жизненного цикла переселяются из организма одного вида в другой). Паразитоиды (как правило, насекомые) на определенных стадиях жизненного цикла ведут свободный образ жизни (питаются как фитофаги или зоофаги). После этого они откладывают яйца в тело организма‑хозяина, в котором паразитируют личинки. Паразитоиды представляют переход к хищникам. Насекомые‑паразитоиды используются для биологического метода контроля насекомых‑вредителей в сельском хозяйстве (трихограмма, теленомус‑наездник и др.).
Эндотрофные и эктотрофные паразиты. Эндотрофные паразиты живут внутри организма‑хозяина (глисты в кишечнике позвоночных, стеблевые нематоды в стеблях хлебных злаков), а эктотрофные – на поверхности организма‑хозяина (блохи, вши, клещи и др.). Особый случай эктотрофного паразитизма – образ жизни карликовых самцов глубоководных удильщиков, которые внедряются острыми зубами в тело самки, после чего тела хозяина и паразита сливаются воедино (объединяются даже кровеносные системы). Тело самки в 13 раз больше тела самца. Экологический смысл этой адаптации заключается в повышении вероятности оплодотворения. Удильщики живут в полной темноте, и иные варианты поиска «спутника жизни» реализовать крайне сложно.
Суперпаразиты – «паразиты паразитов». Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), верхний уровень представлен бактериями или вирусами. Эту матрешку очень точно описал Дж. Свифт:
Под микроскопом он открыл, что на блохе
Живет блоху кусающая блошка;
На блошке той – блошинка‑крошка,
В блошинку же вонзает зуб сердито
Блошиночка… и так ad infinitum.
В этом случае из паразитов формируется пищевая цепь.
Различаются группы паразитов и по сложности жизненного цикла. Одни виды паразитов передаются при непосредственном контакте особей хозяина (например вирусы и бактерии, вызывающие болезни человека). Другие паразиты перед заражением основного хозяина, в теле которого они образуют потомство, проходят через один или несколько видов промежуточных хозяев (например широкий лентец – паразит человека, но в течение жизненного цикла он проходит через стадии жизни в рачках‑циклопах и рыбах).
И, наконец, кроме паразитов, существуют еще и полупаразиты. Значительное число видов растений (в средней полосе в первую очередь из семейства норичниковых) сочетает автотрофное питание и паразитирование на корнях растений‑хозяев. При этом если каждый вид растений‑паразитов (из семейств заразиховых, повиликовых и др.) имеет своего «персонального» хозяина, то число видов‑хозяев для одного вида полупаразита исчисляется десятками и сотнями. В этом случае нет необходимости в тонком подстраивании метаболизма хозяина и полупаразита, как при абсолютном паразитизме, полупаразит получает от растения‑хозяина только неспецифические элементы питания.
Среди полупаразитов есть виды, наносящие вред хозяйству человека. Например погремки (Rhinanthus) при доминировании в луговых сообществах становятся основными продуцентами органического вещества. В этом случае за счет потери энергии при переходе с одного трофического уровня на другой (см. 10.4) биологическая продукция травостоя снижается в 2‑3 раза. Причем погремок не поедается сельскохозяйственными животными ни на пастбище, ни в сене.
В естественных экосистемах взаимоотношения «паразит – хозяин» являются одним из важных факторов поддержания экологического равновесия. Особенно велика их роль при контроле плотности популяций крупных животных, у которых нет естественных врагов‑хищников (слон, бегемот, крокодил, лев и др.). При отсутствии паразитов их отношения с жертвами могли бы быть нарушены.
В процессе длительной коэволюции (взаимоприспособления) паразитов и хозяев вырабатываются специальные механизмы, которые позволяют им устойчиво сосуществовать.
Защитные реакции хозяев могут быть следующими:
– иммунный ответ организма, т.е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов;
– сбрасывание зараженных частей (это особенно характерно для растений‑хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги;
– выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при паразитировании тли);
– изоляция органов поражения как «зеленых островов» (формирование галлов у дуба, орешника и других растений после того, как насекомое‑паразитоид отложит в ткани листа яйцо);
– уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, которые таким образом снижают долю зараженных особей в популяции;
– формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
В естественных экосистемах формирование экологического равновесия между популяциями паразитов и их хозяев – нормальное явление. При этом в отличие от отношений «фитофаг – растение» или «хищник – жертва» оно возможно без третьего участника. В силу того, что паразиты связаны с ограниченным кругом хозяев, эта связь математически описывается проще, чем связь между хищниками и их жертвами. Во многих случаях проявляется модель Лотки–Вольтерры: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев.
Ситуация изменяется в антропогенных экосистемах, особенно в сельскохозяйственных, где заражение скота паразитами может привести к гибели многих животных. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим – лямблией), болезнями бактериальной и вирусной природы.
Катастрофическими бывают последствия заноса паразитов в новые районы, где у их потенциальных хозяев отсутствуют механизмы защиты от паразитов. В ХХ в. произошли ботанические катастрофы в Америке (гибель зубчатого каштана Castanea dentata от занесенного туда из Китая паразитического гриба Endothia parasitica, вызывающего «рак каштана») и Европе, где от «голландской болезни» почти полностью исчез вяз. Болезнь вызывает гриб Ophiostoma ulmi, который переносится жуком‑короедом.
К еще более серьезным последствиям привело распространение вируса «коровьей чумы» в саваннах Африки (McNaughton, 1992). Под влиянием этого вируса, который первоначально вызвал болезни у домашнего скота, а затем поразил многие виды диких крупных копытных животных, резко снизилась нагрузка фитофагов на растительность саванны, и в первую очередь на ее древесно‑кустарниковый компонент и на его отношения с травяным покровом саванны. Кустарники и деревья стали бурно разрастаться, подавляя травы, что увеличило количество пожаров, которые при таком сомкнутом древесном пологе стали более частыми. После пожаров деревья отрастали плохо и замещались кустарниками, корневища которых позволяли им сохраняться во время пожара. Только в 70‑е годы, когда удалось снять влияние на экосистемы вируса «коровьей чумы», вылечив от болезни домашний скот, поставлявший паразита популяциям диких животных через выделение слюны на пастбищах, где их выпас чередовался, процесс был остановлен. Восстановилась плотность популяций животных, питающихся ветками, и соответственно восстанавился баланс между древесно‑кустарниковым и травяным компонентами экосистемы саванны, которая приобрела первозданный облик.
Есть примеры более сложных взаимоотношений «паразит – хозяин» с посредником. Так гетеротрофное растение‑паразит подъельник паразитирует на грибах, разлагающих мертвое органическое вещество, но, кроме того, по гифам микоризного гриба, как по шлангу, выкачивает питательные элементы из корней ели.
Контрольные вопросы
1. Чем паразиты отличаются от хищников?
2. Расскажите о разнообразии паразитов.
3. Какие защитные реакции против паразитов вырабатываются у хозяев?
4. Расскажите о нарушении экологического равновесия в паре «паразит – хозяин» при вмешательстве человека.
|
|
|
|
|
Дата добавления: 2014-12-27; Просмотров: 1151; Нарушение авторских прав?; Мы поможем в написании вашей работы!