КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Этногеномика народов волго-уральского региона
|
|
|
|
В Волго-Уральском регионе столкнулись две волны расселения: европеоидная и монголоидная. Находясь на границе двух частей света - Европы и Азии, этот регион на протяжении исторически длительного времени был местом взаимодействия многих этнических слоев. В формировании народов края известна роль угров Западной Сибири, финнов севера Восточной Европы, индо-иранцев Ближнего Востока, тюрков Южной Сибири и Алтая, а позднее кочевых татаро-монголь-ских племен и славянских народов Центральной и Западной Европы. Вплоть до XVI в. Среднее Поволжье было подлинным "котлом", в котором перемешались многие этнические группы. Следы взаимопроникновения хранят гены проживающих здесь народов.
Современные популяции Волго-Урала крайне неоднородны по этнолингвистической структуре. Сейчас здесь проживают представители финно-угорской ветви Уральской языковой семьи (удмурты, марийцы, мордва, коми), тюркской ветви Алтайской языковой семьи (башкиры, татары, чуваши) и восточно-славянской ветви Индоевропейской языковой семьи (русские). С целью познания факторов формирования генофонда, генетической реконструкции родства и происхождения современных народов Волго-Урала изучены 17 полиморфных аутосомных ДНК-локусов, определена нуклеотидная последовательность ДНК гипервариабельного участка митохондриального генома, выполнен анализ полиморфизма 24 локусов Y-хромосомы. В ходе исследований проанализированы геномы 1500 индивидов из 8 популяций (башкиры, татары, русские, чуваши, удмурты, коми, мордва, марийцы).
Полиморфизм аутосомных ДНК-локусов. Анализ полиморфизма 17 аутосомных ДНК-локусов в популяциях Волго-Уральского региона свидетельствует об их неоднородности и высокой информативности для популяционно-генетических исследований. Оценка степени генного разнообразия (Gst) народов этого региона показала, что их генофонд не только географически занимает промежуточное место между европейскими и сибирскими генофондами. Уровень генетических различий между этносами Волго-Урала (Gst = 1.91%) занимает как бы промежуточное положение между европейскими (Gst = 1.18%) и сибирскими народами (Gst = 5.84%), тяготея к европейскому типу (рис. 2). Изучение соотношения европеоидного и монголоидного вкладов в генофонд народов Волго-Уральского региона по данным полиморфизма ДНК-локусов выявило большую долю европеоидности. Самая высокая доля европеоидности обнаружена в популяциях мордвы (89.4%) и коми (81.0%), самая низкая - в популяциях башкир (62%) и марийцев (53%) [6]. Это согласуется с данными антропологии и археологии.
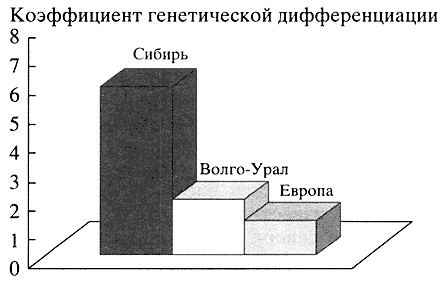
Рис. 2. Уровень генетических различий между этносами Сибири, Волго-Уральского региона и Европы
Полиморфизм мтДНК. Чтобы судить о генетическом разнообразии народов Волго-Уральского региона по материнской линии, была определена последовательность гипервариабельного участка митохондриальной ДНК (377 нуклеотидов) и проведен анализ полиморфизма мтДНК с использованием 26 эндонуклеаз (ферментов рестрикции). Полученные данные анализировались методом так называемых медианных сетей. Медианная сеть - один из графических вариантов представления данных о полиморфизме ДНК, который позволяет проследить дивергенцию митохондриального генома от предковой ДНК. В качестве предкового выбирается гаплотип, который широко распространен в данной популяции и включает наиболее частые аллели, характерные для данного этноса. В процессе такого анализа в единую медианную сеть соединяются посредством линий сотни и тысячи соседствующих, наиболее близких индивидуумов. В этой сети узловые соединения совпадают со специфическими гаплотипами, определенными по характерным мутациям изучаемого генома.
При сравнительном изучении мтДНК у народов Волго-Уральского региона выявленные гап-лотипы были отнесены к определенным линиям в соответствии с классификацией Макуолли [7]. Для большинства расовых и географических групп установлена специфичность типов мтДНК (рис. 3). Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям Н, J, V, 1, К, Т, U, W и X, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера М и N, которые соединяются в африканском макрокластере L3. Последний рассматривается в качестве общего предка для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру М, который подразделяется на линии С, Z, D, G, Е и др. [8]. Суперкластер N в Азии делится на несколько линий: А, Y, В, F.
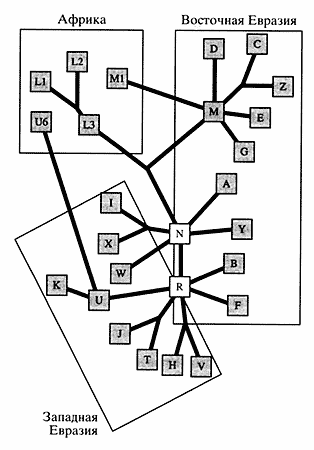
| Рис. 3. Медианная сеть, демонстрирующая полиморфизм мтДНК в популяциях Европы, Азии и Африки Линии D, С, Z, G суперкластера М, линии A, Y суперкластера N и линии В, F суперкластера R характерны для азиатских популяций; линии 1, X, W суперкластера N и линии V, Н. Т, J, U, К суперкластера R - для европейских; линии LI, L2, L3, М1 и U6 принадлежат африканским народам |
Большинство типов мтДНК народов Волго-Уральского региона соответствует линиям мтДНК Европы и Ближнего Востока, что свидетельствует об общих предковых линиях мтДНК, специфичных для европейцев [9]. В целом среди изученных нами популяций частота европейских типов мтДНК оказалась наиболее высокой у мордвы, коми-зырян и русских. С другой стороны, уровень распространения линий мтДНК, специфичных для Восточной Евразии, также достигает больших значений, что ранее не было показано для Западной Европы. Высокая частота линий G, D, С, Z и F в некоторых этнических группах, как тюркских (башкиры), так и финноугорских (удмурты, коми-пермяки), указывает на значительное участие сибирского и центральноазиатского компонента в этногенезе народов Волго-Уральского региона.
Самостоятельный интерес вызывает высокая частота азиатской линии F (6%) у башкир. Эта линия характерна для народов Средней Азии - казахов, уйгуров и монголов, и мы можем предположить, что, во-первых, существенную роль в формировании данной этнической группы сыграл среднеазиатский компонент и, во-вторых, популяция башкир длительное время находится в изоляции от своих ближайших соседей. У других тюркоязычных и финноугорских популяций Волго-Уральского региона частота азиатских линий низкая. Поскольку очевидных географических барьеров как между тюркскими этносами, так и между финноугорскими популяциями не существует, можно сделать вывод о различной демографической истории башкир по материнской линии внутри упомянутых языковых семей. Выборки башкир и удмуртов по совокупности всех полученных по материнской линии данных можно охарактеризовать как этносы, имеющие в прошлом период резкого роста численности в условиях относительной изоляции. Анализ митохондриального генома татар, чувашей, марийцев, мордвы, коми и русских, скорее всего, отражает процессы продолжающейся интенсивной метисации в условиях сохранения постоянного популяционного размера. В целом, медианные сети показывают смешение и взаимопроникновение гаплотипов мтДНК, что свидетельствует как о тесных этногенетических контактах изученных этносов, так и о единой генетической основе населения Волго-Уральского региона по материнской линии.
В распространении типов мтДНК у народов Волго-Уральского региона ведущую роль играют факторы этнокультурной и территориальной близости или отдаленности, но не лингвистические барьеры. Это означает, что по материнской линии финноугорские народы имеют большее сходство с их непосредственными тюркскими соседями, чем с родственными в языковом отношении балтофинскими народами.
Анализ европеоидного и монголоидного вклада в материнские генетические линии народов Волго-Уральского региона не выявил корреляции языка и геномного состава этнических групп. На языках тюркской группы, привнесенных из Азии, говорят не только башкиры (65% монголоидности), но и татары и чуваши, у которых преобладает европеоидный генетический компонент. В остальных популяциях региона вклад монголоидного компонента составляет от 12% у русских до 20% у удмуртов. Русские, проживающие на территории этого региона, имеют 10-12% монголоидных типов мтДНК, а русские из Рязанской и Курской областей - только 2-3%. Это можно объяснить смешением русских с тюркоязычными народами на территории Волго-Уральского региона.
Интересно, что некоторые материнские линии у разных народов, например, у русских, татар и марийцев, оказались общими. Это показывает глубокое родство народов, говорящих на разных языках, придерживающихся разных религий и традиций.
Сравнительный анализ типов мтДНК в 18 популяциях Евразии, включая популяции Волго-Уральского региона (гагаузов, турков, татар, башкир, чувашей, карачаевцев, кумыков, азербайджанцев, узбеков, казахов, киргизов, ногайцев, уйгуров, шорцев, тувинцев, долган, якутов), которые относятся к тюркской ветви Алтайской языковой семьи, позволил установить западно-восточный градиент увеличения частоты азиатских линий мтДНК на расстоянии 8000 км: от 1% у гагаузов из Молдавии до 95% у якутов и 99% у долган (рис. 4). Кроме того, установлено, что лингвистическое сходство популяций играет меньшую роль, чем географическая близость или отдаленность популяций.
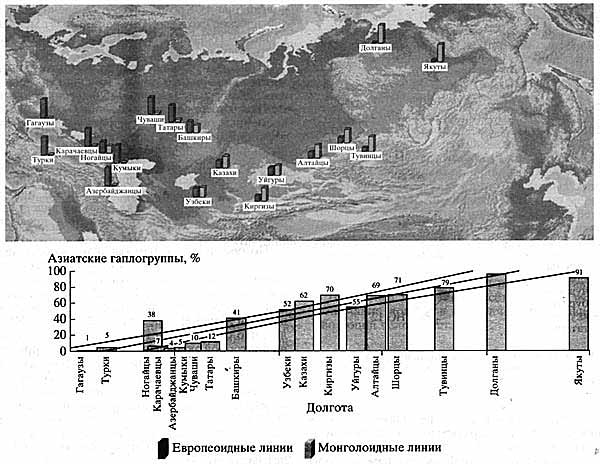
Рис. 4. Результаты сравнительного анализа типов мтДНК в 18 популяциях Евразии
Отчетливо виден западно-восточный градиент увеличения частоты азиатских линий мтДНК
Один из наиболее важных аспектов анализа митохондриального генома - оценка времени коалесценции (расхождения, дивергенции) линий мтДНК в пределах каждой линии. Безусловно, на временные оценки будут влиять различные факторы формирования разнообразия мтДНК: объем выборки, миграция населения, резкий рост численности, феномен "бутылочного горлышка" - сильное сокращение численности наших предков, вызванное, по-видимому, изменением климата, и т.д. Тем не менее оценка времени дивергенции линии возможна при обнаружении предковых гаплотипов.
По ориентировочным оценкам, возраст дивергенции линий, выявленных у народов Волго-Уральского региона, варьировал от 273 ± 57 тыс. лет для азиатской линии Z до 22.76 ± 5.250 тыс. лет для линии С. Возраст дивергенции самой крупной европейской линии Н определен в 20.036 ± 4.250 тыс. лет, что соответствует археологическому времени повторной экспансии населения на территории Урала в постледниковый период. Используя данные о числе мутационных замен и скорости накопления мутации для гипервариабельного участка мтДНК, равной одной мутационной замене за 20.18 тыс. лет, мы получили среднее значение времени дивергенеции мтДНК для народов Волго-Уральского региона. Оно составляет 49.60 тыс. лет назад, что соответствует периоду расселения человека на европейском континенте в эпоху верхнего палеолита.
Полиморфизм ДНК Y-хромосомы. Анализ Y-хромосомы вошел в арсенал методов эволюционной генетики лишь в самое последнее время, когда был найден ряд высокоинформативных полиморфных локусов в ее нерекомбинантной части. Генетические свойства Y-хромосомы, такие как передача только по отцовской линии, отсутствие рекомбинации, малая эффективная численность пула Y-хромосом по сравнению с аутосомами (в четыре раза меньше, чем у аутосом), позволяют прослеживать по гаплотипам Y-хромосомы отцовские линии, представляющие собой последовательную "запись" мутаций в ряду поколений. По сравнению с митохондриальным геномом, насчитывающим 16.5 тыс. пар нуклеотидов. Y-хромосома, размер которой оценивается приблизительно в 60 млн. пар нуклеотидов, дает в руки исследователей потенциально более мощное "оружие".
Если предшествующие работы, посвященные анализу Y-хромосомы в популяциях России, основывались преимущественно на анализе 9 маркеров, то для изучения и сравнения генетического разнообразия отцовских линий в популяциях Волго-Уральского региона использовались 24 маркера Y-хромосомы. В качестве примера на рисунке 5 представлена медианная сеть линий 12 и 16 Y-хромосомы как наиболее интересных в контексте финноугорских народов. Линия 16 практически отсутствует в западноевропейских популяциях, но частота ее высокая среди народов Балтии - эстонцев и финнов, а также у народов Волго-Уральского региона, особенно удмуртов и коми-зырян [10].
| Рис. 5. Медианная сеть линий HG12 и HG16 Y-хромосомы, построенная для некоторых популяций Европы и Азии | 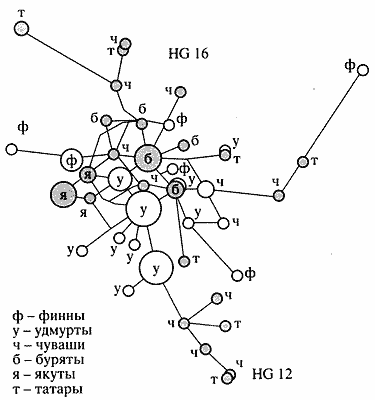
|
Для линии 16 Y-хромосомы уровень генетического разнообразия намного выше в популяциях Восточной Европы (чуваши, татары), чем в изученных популяциях Сибири. Хотя удмурты и имеют очень высокую частоту линий 12 и 16, уровень генетического разнообразия у них невысокий по сравнению с другими популяциями Европы. Данные о низком уровне генетического разнообразия удмуртов получены и по материнской линии при анализе полиморфизма митохондриальной ДНК. Все это свидетельствует о несомненной роли эффекта основателя и дрейфа генов в демографической истории удмуртов.
Анализ распространения и разнообразия линии 16 Y-хромосомы среди восточноевропейских популяций показывает, что местом ее "рождения", возможно, является Восточно-Европейская равнина. Согласно филогеографическому анализу этой линии в Евразии, она начала распространяться с запада на восток. В то же время частота распространения линии 12 - предковой для линии 16 - меньше, чем для линии 16.
Особенно характерна для популяций Волго-Уральского региона линия 3 Y-хромосомы, частота встречаемости которой максимальна у славян (русские и поляки), а также у населения Латвии, Литвы и Эстонии [10]. Таким образом, носители этой линии - этнические группы, принадлежащие к разным языковым семьям. Частота встречаемости таких групп уменьшается в направлении север (Финляндия, Швеция) - юг (Турция, Кавказ). Анализ распространения данной линии в популяциях Волго-Уральского региона подтверждает гипотезу о возможном движении населения после Ледникового периода (Last Glacial Maximum) с территории нынешней Украины, где в то время располагался один из центров потепления [1].
Судя по результатам анализа отцовских линий в популяциях Восточной Европы и, в частности, Волго-Уральского региона, главную роль в формировании генетического разнообразия народов, проживающих на этой территории, по-видимому, играет географическая близость, а не языковая принадлежность. И хотя многие особенности генетической близости популяций объясняются с точки зрения их географического положения, в некоторых случаях "индивидуальная" демографическая история популяции имеет существенное значение. Хороший пример - популяция удмуртов, у которых разнообразие линий Y-хромосомы и мтДНК ограничено. Принимая темпы мутирования изученных ДНК-маркеров Y-хромосомы за 2.1 х 10-3 и длительность одного поколения за 25 лет, получаем, что наблюдаемая дисперсия гаплотипов, выявленных у современного населения Волго-Уральского региона, сформировалась приблизительно 42.5 тыс. лет назад, что соответствует времени заселения Европы человеком в эпоху верхнего палеолита.
Таким образом, исследования полиморфизма аутосомных, митохондриальных и Y-хромосомных ДНК-маркеров внесли важный вклад в понимание путей происхождения человека и рас, расселения Homo sapiens по планете, в генетическую и демографическую историю отдельных этносов и популяций. Можно надеяться, что по мере все более подробного изучения свойств конкретных маркеров ДНК будут появляться дополнительные возможности для изучения генетической истории народов Европы и Азии. Дальнейшее развитие этногеномики в сочетании с палео- и археогеномикой значительно расширит наши представления о генофонде человека, внесет весомый вклад в понимание вопросов исторического развития и эволюции человечества.
|
|
|
|
|
Дата добавления: 2015-04-24; Просмотров: 2912; Нарушение авторских прав?; Мы поможем в написании вашей работы!