КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транспозоны
|
|
|
|
Инсерционные последовательности
Простейшим типом мобильных элементов являются вставочные или инсерционные последовательности (IS – элементы) (англ. insertion —вставка, sequence — последовательность), несущие только один ген транспозазы, с помощью которой IS–элементы могут встраиваться в различные участки хромосомы. Это подвижные фрагменты ДНК размером до 1500 пар нуклеотидов, встречаются в ДНК всех организмов.
Впервые они были открыты у бактерий при обнаружении вставок (инсерций) нового материала в пределах бактериальных оперонов. Такие вставки локализовались внутри гена и предотвращали его транскрипцию. Восстановление активности гена происходило лишь после утраты встроенного материала. При сравнении последовательностей, присутствующих в различных инсерционных мутантах, выявлено несколько типов IS–элементов, хотя определенные черты являлись общими для всех элементов. Каждый из них представляет собой автономную единицу, кодирующую белок (или белки), необходимый для транспозиции (перемещения). Этот белок узнает концы транспозирующегося элемента, которые образованы инвертированными повторяющимися последовательностями разной протяженности. Если две копии одного и того же элемента находятся в хромосоме на относительно небольшом расстоянии друг от друга, они могут транспозироваться в виде одной генетической структуры: два IS–элемента и заключенные между ними бактериальные гены.
Отличие IS – последовательностей от транспозонов: не имеют в своем составе структурных генов, а только гены, отвечающие за перемещение (транспозицию) в различные участки ДНК.
Пути перемещения IS – последовательностей:
– консервативный — покидая один участок, IS–элемент встраивается в другой;
– репликативный — синтезируется копия, которая встраивается в другой участок генома.
Состояние IS – элементов в бактериальной клетке: не обнаружены в свободном состоянии (не способны к автономной репликации).
Биологическая роль IS – элементов:
– координирующая взаимодействие внехромосомных факторов наследственности (плазмид, умеренных фагов, транспозонов) между собой и с бактериальной хромосомой для обеспечения их рекомбинации;
– регуляторная — участвуют в регуляции активности (инактивации или активации) генов бактериальной клетки, в которых произошла вставка IS–элемента;
– индукция мутаций при включении IS–элементов в бактериальную хромосому;
– являются генетическими маркерами вида (рода) бактерий.
Транспозоны (Tn–элементы) — более сложно организованные транспозирующиеся и самоинтегрирующиеся фрагменты ДНК длиной 2 до 25 тысяч пар нуклеотидов. Способны менять место своей локализации в молекуле ДНК, а также мигрировать из одной молекулы ДНК в другую. Транспозоны распространяются среди различных видов бактерий, встраиваясь и перемещаясь среди хромосом, плазмид, умеренных фагов.
Транспозоны имеют особые концевые структуры нескольких типов, позволяющие отличать их от других фрагментов ДНК. Это позволило обнаружить транспозоны не только у бактерий и дрожжей, но и в клетках растений, насекомых, позвоночных животных и человека. При интеграции транспозонов в хромосому клеток животных или человека они приобретают сходство с провирусами.
Состояние транспозонов в бактериальной клетке:
– интегрированное в репликон (реплицируется вместе с ним); при включении в бактериальную ДНК транспозоны вызывают в ней дупликации, а при перемещении — делеции и инверсии,
– свободное автономное (замыкается в кольцо и не реплицируется).
Таким образом, транспозоны, как IS–последовательности, не способны к самостоятельной репликации и размножаются только в составе бактериальной хромосомы.
Отличие транспозонов от IS – последовательностей: содержат в своем составе не только гены транспозиции, но и структурные гены.
Состав транспозонов:
– один или несколько специфических структурных генов, детерминирующих синтез молекул со специфическими биологическими свойствами: токсинообразование, синтез ферментов (напр., гемолизинов, лактазы), резистентность к антибиотикам, устойчивость к солям тяжелых металлов,
– гены транспозиции: два IS–элемента—концевые структуры, отличающие транспозон от других фрагментов ДНК.
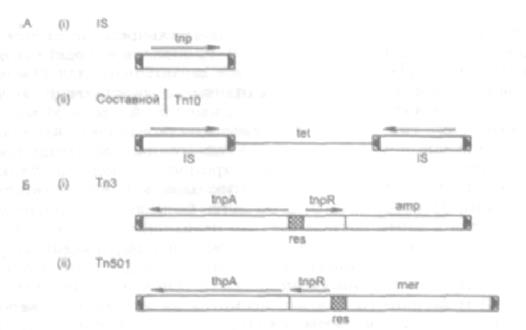
Рис. 73. Классы бактериальных транспозонов:
А — инсерционный элемент (IS) (i); составной транспозон (ii) — (Tn10);
Б — Tn3 и Tn501 — транспозоны семейства Tn3.
Стрелки на концах транспозонов — короткие инвертированные последовательности, узнаваемые ферментом траспозазой (ген tpn). tet, amp и mer — гены в составе транспозонов, кодирующие резистентность к тетрациклину, ампициллину и солям ртути соответственно.
По особенностям строения различают (рис. 73):
– составные транспозоны —в их состав входят два полных IS–элемента, фланкирующих центральную часть, которая содержит гены резистентности к антибиотикам или гены термолабильного токсина. IS–элементы содержат информацию о способности к транспозиции. У некоторых транспозонов этого типа такая информация сохраняется только в одном из IS–элементов, а во втором гены, кодирующие эту информацию, инактивированы мутациями. На концах IS–элементов имеются короткие инвертированные повторяющиеся последовательности, имеющие важное значение для транспозиции, так как именно они узнаются ферментом транспозазой, осуществляющим транспозицию;
– транспозоны ТnЗ – семейства —не содержат в своем составе IS–элементов, а ограничены высоко гомологичными инвертированными концевыми повторами, состоящими из 35–48 пар нуклеотидов; центральные области этих транспозонов содержат информацию об их транспозиции, а также гены, не связанные с транспозицией (напр., кодирующие резистентность к антибиотикам или к ртути).
Пути перемещения транспозонов:
– консервативный — транспозон вырезается из одного участка и перемещается в другой без увеличения количества копий; при этом участок ДНК, откуда вырезается транспозон, утрачивает свою функцию,
– репликативный — синтезированная копия транспозона перемещается в новое место, при этом увеличивается количество копий. Новые копии транспозонов могут мигрировать в плазмиды и ДНК фагов, которые в свою очередь, проникая в бактериальные клетки, способствуют их распространению в популяции.
Биологическая роль транспозонов:
– регуляторная — участвуют в регуляции активности (инактивации или активации) генов. Обеспечивая перенос генов внутри клетки, способствуют их интеграции в плазмиды и бактериофаги и распространению в популяциях клеток разных видов, придавая им новые свойства, часто связанные со способностью выживать в неблагоприятных условиях. Интеграция транспозонов может привести к экспрессии соседнего «молчащего» гена;
– индукция геномных мутаций разного типа;
– кодирующая — осуществляют горизонтальный перенос генов несущих информацию о синтезе токсинов, ферментов. Некоторые транспозоны приобретают новые гены резистентности к антибиотикам, существующие в виде генных циркулярно-замкнутых кассет. Это происходит, когда в составе транспозона содержатся дополнительные генетические структуры —интегроны,отвечающие за сайт-специфическую рекомбинацию. Интегроны содержат ген интегразы, промотор и сайт интеграции. С помощью фермента интегразы захватывают генные кассеты, внедряют их в специфический сайт интегрона и экспрессируют их со своего промотора;
– являются генетическими маркерами вида (рода) бактерий.
Сравнительная характеристика внехромосомных факторов наследственности прведена в табл. 36.
Таблица 36
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 4837; Нарушение авторских прав?; Мы поможем в написании вашей работы!