КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Побег и системы побегов
|
|
|
|
ЛЕКЦІЯ
Корінь як орган рослини. Функції коренів, види, типи кореневих систем. Зони кореня. Спеціалізація та метаморфози коренів.
Пагін як орган рослини. Функції, частини пагону. Різноманітність стебел за положенням у просторі, формою поперечного перерізу тощо. Визначення бруньки та її будова, класифікація за місцем розташування, структурою, функціями. Метаморфози стебел.
Листок як орган рослини. Частини листків, особливості їх будови та функції. Типи листків та їх морфологічна різноманітність. Розміщення листків. Метаморфози листка.
Значення й використання вегетативних органів рослин.
Корень – основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного питания. Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не образуются листья, а апикальная меристема всегда прикрыта корневым чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции:
1) корни укрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вынесение побегов наверх;
2) в корнях синтезируются различные вещества, которые затем передвигаются в другие органы растения;
3) в корнях могут откладываться запасные вещества;
4) корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве.
Совокупность корней одной особи образует единую в морфологическом и физиологическом отношении корневую систему.
В состав корневых систем входят корни различной морфологической природы – главный корень, боковые и придаточные корни.
Главный корень развивается из зародышевого корешка. Боковые корни образуются на корне (главном, боковом, придаточном), который по отношению к ним обозначается как материнский. Они возникают на некотором расстоянии от апекса, в направлении от основания корня к его верхушке. Боковые корни закладываются эндогенно, т.е. во внутренних тканях материнского корня. Если бы ветвление происходило в самом апексе, это бы затруднило продвижение корня в почве. Придаточные корни могут возникать и на стеблях, и на листьях, и на корнях.
По происхождению выделяют следующие типы корневых систем:
1) система главного корня(стержневая) представлена главным корнем (первого порядка) с боковыми корнями второго и последующих порядков (у многих кустарников и деревьев, большинства двудольных растений);
2) система придаточных корней(мочковатая) развивается на стеблях, листьях; встречается у большинства однодольных растений и многих двудольных, размножающихся вегетативно;
3) смешанная корневая система образована главным и придаточными корнями с их боковыми ответвлениями (многие травянистые двудольные).
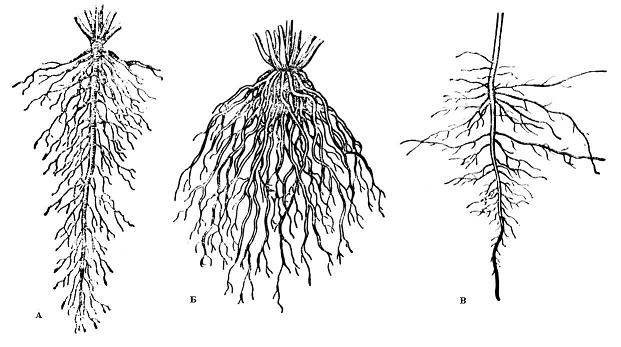
Рис. 4.1. Типы корневых систем: А – система главного корня; Б – система придаточных корней; В – смешанная корневая система (А и В – стержневые корневые системы; Б – мочковатая корневая система).
По форме различают стержневую и мочковатую корневые системы.
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди остальных корней. В мочковатой корневой системе главный корень незаметен или его нет, а корневая система составлена многочисленными придаточными корнями (рис. 4.1).
Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Чехлик состоит из живых клеток и постоянно обновляется: по мере того, как с его поверхности слущиваются старые клетки, на смену им, изнутри, апикальная меристема образует новые молодые клетки. Наружные клетки корневого чехлика отслаиваются еще будучи живыми, они продуцируют обильную слизь, которая облегчает продвижение корня среди твердых частиц почвы. В клетках центральной части чехлика содержится много крахмальных зерен. По-видимому, эти зерна служат статолитами, т. е. способны перемещаться в клетке при изменении положения кончика корня в пространстве, благодаря чему корень растет всегда в сторону действия силы тяжести (положительный геотропизм).
Под чехликом находится зона деления, представленная апикальной меристемой, в результате деятельности которой формируются все прочие зоны и ткани корня. Зона деления имеет размеры около 1 мм. Клетки апикальной меристемы относительно мелкие, многогранные, с густой цитоплазмой и крупным ядром.
Вслед за зоной деления располагается зона растяжения, или зона роста. В этой зоне клетки почти не делятся, а сильно растягиваются (растут) в продольном направлении, вдоль оси корня. Объем клеток увеличивается за счет поглощения воды и образования крупных вакуолей, при этом высокое тургорное давление проталкивает растущий корень между частицами почвы. Протяженность зоны растяжения обычно невелика и не превышает нескольких миллиметров.
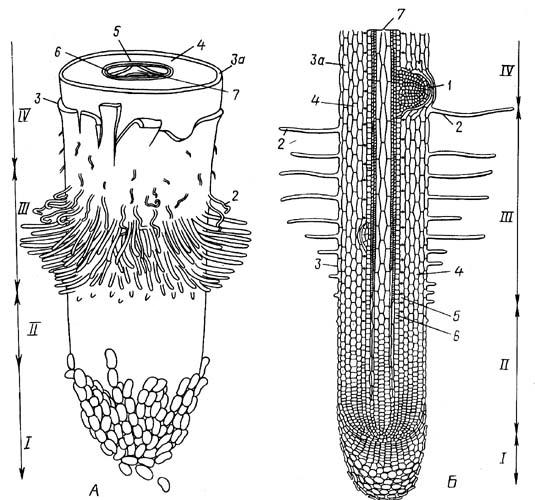
Рис. 4.2. Общий вид (А) и продольный срез (Б) корневого окончания (схема): I – корневой чехлик; II – зоны деления и растяжения; III – зона всасывания; IV – начало зоны проведения: 1 – растущий боковой корень; 2 – корневые волоски; 3 – ризодерма; 3а – экзодерма; 4 – первичная кора; 5 – эндодерма; 6 – перицикл; 7 – осевой цилиндр.
Далее идет зона поглощения, или зона всасывания. В этой зоне покровной тканью является ризодерма (эпиблема), клетки которой несут многочисленные корневые волоски. Растяжение корня прекращается, корневые волоски плотно охватывают частицы почвы и как бы срастаются с ними, поглощая воду и растворенные в ней минеральные соли. Зона поглощения имеет протяжение до нескольких сантиметров. Эту зону называют также зоной дифференциации, поскольку именно здесь происходит образование постоянных первичных тканей.
Продолжительность жизни корневого волоска не превышает 10-20 дней. Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие органы растения. В зоне проведения формируются боковые корни (рис. 4.2).
Специализация и метаморфозы корней. У большинства растений в одной и той же корневой системе отчетливо различаются ростовые и сосущие окончания. Ростовыеокончания обычно более мощные, быстро удлиняются и продвигаются вглубь почвы. Зона растяжения у них хорошо выражена, и апикальные меристемы работают энергично. Сосущие окончания, возникающие в большом количестве на ростовых корнях, удлиняются медленно, и их апикальные меристемы почти перестают работать. Сосущие окончания как бы останавливаются в почве и интенсивно ее «обсасывают».
У древесных растений различают толстые скелетные и полускелетные корни, на которых образуются недолговечные корневые мочки. В состав корневых мочек, непрерывно заменяющих друг друга, входят ростовые и сосущие окончания.
Если корни выполняют особые функции, их строение меняется. Резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций, носит название метаморфоза. Видоизменения корней очень разнообразны.
Корни многих растений образуют симбиоз с гифами почвенных грибов, называемый микоризой («грибокорень»). Микориза образуется на сосущих корнях в зоне поглощения. Грибной компонент облегчает корням получение воды и минеральных элементов из почвы, часто гифы грибов заменяют корневые волоски. В свою очередь, гриб получает от растения углеводы и другие питательные вещества. Различают два основных типа микоризы. Гифы эктотрофной микоризы образуют чехол, окутывающий корень снаружи. Эктомикориза широко распространена у деревьев и кустарников. Эндотрофная микориза встречается в основном у травянистых растений. Эндомикориза находится внутри корня, гифы внедряются в клетки коровой паренхимы. Микотрофное питание очень широко распространено. Некоторые растения, например орхидные, вообще не могут существовать без симбиоза с грибами.
На корнях бобовых возникают особые образования – клубеньки, в которых поселяются бактерии из рода Rhizobium. Эти микроорганизмы способны усваивать атмосферный молекулярный азот, переводя его в связанное состояние. Часть веществ, синтезированных в клубеньках, усваивают растения, бактерии, в свою очередь, используют вещества, находящиеся в корнях. Этот симбиоз имеет большое значение для сельского хозяйства. Бобовые растения благодаря дополнительному источнику азота богаты белками. Они дают ценные пищевые и кормовые продукты и обогащают почву азотистыми веществами.
Очень широко распространены запасающие корни. Они обычно утолщены и сильно паренхиматизированы. Сильно утолщенные придаточные корни называют корневыми шишками, или корнеклубнями (георгин, некоторые орхидные). У многих, чаще двулетних, растений со стержневой корневой системой возникает образование, носящее название корнеплода. В образовании корнеплода принимают участие и главный корень, и нижняя часть стебля. У моркови почти весь корнеплод составлен корнем, у репы корень образует лишь самую нижнюю часть корнеплода (рис. 4.12). Корнеплоды культурных растений возникли в результате длительного отбора. В корнеплодах сильно развита запасающая паренхима и исчезли механические ткани. У моркови, петрушки и других зонтичных паренхима сильно развита во флоэме; у репы, редьки и других крестоцветных – в ксилеме. У свеклы запасные вещества откладываются в паренхиме, образованной деятельностью нескольких добавочных слоев камбия (рис. 4.12).
У многих луковичных и корневищных растений образуются втягивающие, или контрактильные корни (рис. 4.13, 1). Они могут укорачиваться и втягивать побег в почву на оптимальную глубину на время летней засухи или зимних морозов. Втягивающие корни имеют утолщенные основания с поперечной морщинистостью. Воздушные корни образуются у многих тропических эпифитных однодольных из семейств орхидных, ароидных, бромелиевых (рис. 4.13, 3). Эпифиты поселяются на других растениях, но не паразитируют на них, а используют как подпорку для поднятия вверх, к свету. Воздушные корни свободно висят в воздухе и приспособлены к поглощению атмосферной влаги. На их поверхности образуется веламен (рис. 4.14). Веламен, как и ризодерма, образуется из протодермы, но это многослойная ткань. Клетки веламена отмирают, их стенки имеют сетчатые или спиральные утолщения. Через поры и сквозные отверстия в клеточных стенках влага проникает капиллярным путем. Изнутри веламен подстилается экзодермой со сложно построенными пропускными клетками, через которые вода передается в клетки коры и осевого цилиндра (рис. 4.14). Веламен встречается и у некоторых наземных однодольных (кливия, хлорофитум), в этом случае он выполняет функцию механической защиты и предохраняет растение от потери воды из первичной коры.
Дыхательные корни, или пневматофоры (рис. 4.13, 2) образуются у некоторых тропических древесных растений, живущих в условиях недостатка кислорода (таксодиум, или болотный кипарис; растения мангровых зарослей, обитающие по болотистым берегам океанических побережий). Пневматофоры растут вертикально вверх и высовываются над поверхностью почвы. Через систему отверстий в этих корнях, связанных с аэренхимой, воздух поступает в подводные органы.
У некоторых растений для поддержания побегов в воздушной среде образуются дополнительные опорные корни. Они отходят от горизонтальных ветвей кроны и, достигнув поверхности почвы, интенсивно ветвятся, превращаясь в столбовидные образования, поддерживающие крону дерева (столбовидные корни баньяна) (рис. 4.15, 2). Ходульные корни отходят от нижних участков стебля, придавая стеблю устойчивость. Они образуются у растений мангровых зарослей, растительных сообществ, развивающихся на затопляемых во время прилива тропических берегах океанов (рис. 4.15, 3), а также у кукурузы (рис. 4.15, 1). У фикуса каучуконосного образуются досковидные корни. В отличие от столбовидных и ходульных, они являются по происхождению не придаточными, а боковыми корнями.
Побег, как и корень, - основной орган растения. Вегетативные побеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные, обеспечивающие размножение.
Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля, имеющего цилиндрическую форму, и листьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являются почки – зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции.
Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел. Участки стебля между соседними узлами – междоузлия. Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеет метамерное строение, метамером (повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие
Первый побег растения – его главный побег, или побег первого порядка. Он образуется из зародышевого побега, заканчивающегося почечкой, которая формирует все последующие метамеры главного побега. По положению эта почка – верхушечная; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров. Кроме верхушечной, на побеге образуются боковые почки. У семенных растений они находятся в пазухах листьев и называются пазушными. Из боковых пазушных почек развиваются боковые побеги, и происходит ветвление, за счет которого увеличивается общая фотосинтезирующая поверхность растения. Формируется система побегов, представленная главным побегом (побегом первого порядка) и боковыми (побегами второго порядка), а при повторении ветвления – боковыми побегами третьего, четвертого и последующих порядков. Побег любого порядка имеет свою верхушечную почку и способен к нарастанию в длину.
^ Почка – это зачаточный, еще не развернувшийся побег. Внутри почки заключена меристематическая верхушка побега – его апекс (рис. 4.17). Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех органов и первичных тканей побега. Источником постоянного самообновления апекса являются инициальные клетки апикальной меристемы, сосредоточенные на кончике апекса. Вегетативный апекс побега, в отличие от всегда гладкого апекса корня, регулярно формирует на поверхности выступы, представляющие собой зачатки листьев. Гладким остается лишь самый кончик апекса, который называют конусом нарастания побега. Его форма сильно варьирует у разных растений и не всегда имеет вид конуса, вершинная часть апекса может быть низкой, полушаровидной, плоской или даже вогнутой.
Из вегетативных почек развиваются вегетативные побеги, состоящие из стебля, листьев и почек. Такая почка состоит из меристематической зачаточной оси, оканчивающейся конусом нарастания, и зачаточных листьев разного возраста. Нижние листовые зачатки вследствие неравномерного роста загибаются внутрь и прикрывают верхние, более молодые, зачатки листьев и конус нарастания. Узлы в почке сближены, так как междоузлия еще не успели вытянуться. В пазухах листовых зачатков в почке уже могут быть заложены зачатки пазушных почек следующего порядка (рис. 4.17). В вегетативно-генеративных почках заложен ряд вегетативных метамеров, а конус нарастания превращен в зачаточный цветок или соцветие. Генеративные, или цветочные почки заключают в себе только зачаток соцветия или одиночного цветка, в последнем случае почку называют бутоном.
Кроме обычных, экзогенных по заложению, пазушных почек у растений часто образуются придаточные, или адвентивные почки. Они возникают не в меристематической верхушке побега, а на взрослой, уже дифференцированной части органа эндогенно, из внутренних тканей. Придаточные почки могут образоваться на стеблях (тогда они обычно расположены в междоузлиях), листьях и корнях. Придаточные почки имеют большое биологическое значение: они обеспечивают активное вегетативное возобновление и размножение тех многолетних растений, у которых они имеются. В частности, при помощи придаточных почек возобновляются и размножаются корнеотпрысковые растения (малина, осина, осот, одуванчик). Корневые отпрыски – это побеги, развившиеся из придаточных почек на корнях. Придаточные почки на листьях образуются относительно редко. Если такие почки сразу дают маленькие побеги с придаточными корешками, которые отпадают от материнского листа и вырастают в новые особи, их называют выводковыми (бриофиллум).
Направление роста побегов. Побеги, растущие вертикально, перпендикулярно поверхности земли, носят название ортотропных. Горизонтально растущие побеги называются плагиотропными. Направление роста может меняться в процессе развития побега.
В зависимости от положения в пространстве различают морфологические типы побегов (рис. 4.19). Главный побег в большинстве случаев сохраняет ортотропный рост и остается прямостоячим. Боковые побеги могут расти в различном направлении, часто они образуют разной величины угол с материнским побегом. В процессе роста побег может изменить направление с плагиотропного на ортотропное, тогда он называется приподнимающимся, или восходящим. Побеги с сохраняющимся в течение жизни плагиотропным ростом называются стелющимися. Если они образуют в узлах придаточные корни, их называют ползучими.
Ортотропный рост связан определенным образом со степенью развития механических тканей. При отсутствии хорошо развитых механических тканей в удлиненных побегах ортотропный рост невозможен. Но нередко растения, не обладающие достаточно развитым внутренним скелетом, все же растут вверх. Это достигается различными путями. Слабые побеги таких растений - лиан закручиваются вокруг какой-либо твердой опоры (вьющиеся побеги), лазят с помощью различного рода шипиков, крючков, корней - прицепок (лазящие побеги), цепляются с помощью усиков различного происхождения (цепляющиеся побеги).
Листорасположение. Листорасположение, или филлотаксис – порядок размещения листьев на оси побега. Различают несколько основных типов листорасположения (рис. 4.20).
Спиральное, или очередное листорасположение наблюдается, когда на каждом узле расположен один лист, и основания последовательных листьев можно соединить условной спиральной линией. Двурядное листорасположение можно рассматривать как частный случай спирального. При этом на каждом узле находится один лист, охватывающий широким основанием всю или почти всю окружность оси. Мутовчатое листорасположение возникает, если на одном узле закладывается несколько листьев. Супротивное листорасположение – частный случай мутовчатого, когда на одном узле образуются два листа, точно друг против друга; чаще всего такое листорасположение бывает накрест супротивным, т.е. соседние пары листьев находятся во взаимно перпендикулярных плоскостях (рис. 4.20).
^ Типы ветвления побега. Ветвление – это образование системы осей. Оно обеспечивает увеличение общей площади соприкосновения тела растения с воздушной средой, водой или почвой. Ветвление возникло в процессе эволюции еще до появления органов. В простейшем случае верхушка главной оси вильчато ветвится и дает начало двум осям следующего порядка. Это верхушечное, или дихотомическое ветвление. Верхушечным ветвлением обладают многие многоклеточные водоросли, а также некоторые примитивные растения, например плауны (рис. 4.21).
Для остальных групп растений характерен более специализированный боковой тип ветвления. При этом боковые ветви закладываются ниже верхушки главной оси, не затрагивая ее способности к дальнейшему нарастанию. При таком способе потенциальные возможности ветвления и образования систем органов гораздо более обширны и биологически выгодны. Различают два типа бокового ветвления: моноподиальное и симподиальное (рис. 4.21). При моноподиальной системе ветвления каждая ось представляет собой моноподий, т.е. результат работы одной верхушечной меристемы. Моноподиальное ветвление характерно для большинства голосеменных и многих травянистых покрытосеменных растений. Большинство покрытосеменных растений, однако, ветвится по симподиальному типу. При симподиальном ветвлении верхушечная почка побега на определенном этапе отмирает или прекращает активный рост, но начинается усиленное развитие одной или нескольких боковых почек. Из них образуются побеги, замещающие побег, прекративший рост. Образовавшаяся при этом ось представляет собой симподий – составную ось, состоящую из осей нескольких последовательных порядков. Способность растений к симподиальному ветвлению имеет большое биологическое значение. В случае повреждения верхушечной почки рост оси будет продолжен боковыми побегами.
Наиболее часто встречающимся подземным метаморфозом побега является корневище (рис. 4.24). Корневищем принято называть долговечный подземный побег, выполняющий функции отложения запасных питательных веществ, возобновления, а иногда и вегетативного размножения. Корневище формируется у многолетних растений, как правило, не имеющих во взрослом состоянии главного корня. По положению в пространстве оно может быть горизонтальным, косым или вертикальным. Корневище обычно не несет зеленых листьев, но, будучи побегом, сохраняет метамерную структуру. Узлы выделяются либо по листовым рубцам и остаткам сухих листьев, либо по живым чешуевидным листьям, также в узлах располагаются пазушные почки. По этим признакам корневище легко отличить от корня. Как правило, на корневище формируются придаточные корни; из почек вырастают боковые ответвления корневища и надземные побеги.
Формируется корневище или изначально как подземный орган (купена, вороний глаз, ландыш, черника), или сначала как надземный ассимилирующий побег, который затем погружается в почву с помощью втягивающих корней (земляника, медуница, манжетка). Корневища могут нарастать и ветвиться моноподиально (манжетка, вороний глаз) или симподиально (купена, медуница). В зависимости от длины междоузлий и интенсивности нарастания, различают длинные и короткие корневища и, соответственно, длиннокорневищные и короткокорневищные растения.
Близки к корневищам подземные столоны – недолговечные тонкие подземные побеги, несущие недоразвитые чешуевидные листья. Столоны служат для вегетативного размножения, расселения и захвата территории. Запасные питательные вещества в них не откладываются.
У некоторых растений (картофель, земляная груша) к концу лета из верхушечных почек столонов образуются клубни (рис. 4.24). Клубень имеет шаровидную или овальную форму, стебель сильно утолщен, в нем откладываются запасные питательные вещества, листья редуцируются, в их пазухах формируются почки. Столоны отмирают и разрушаются, клубни перезимовывают, и на следующий год дают начало новым надземным побегам.
Клубни не всегда развиваются на столонах. У некоторых многолетних растений клубневидно разрастается и утолщается основание главного побега (цикламен, капуста кольраби) (рис. 4.24). Функции клубня – запас питательных веществ, переживание неблагоприятного периода года, вегетативное возобновление и размножение.
Луковица – это, как правило, подземный побег с очень коротким уплощенным стеблем – донцем и чешуевидными мясистыми сочными листьями, запасающими воду и растворимые питательные вещества, главным образом сахара. Из верхушечной и пазушных почек луковиц вырастают надземные побеги, на донце образуются придаточные корни (рис. 4.24). Таким образом, луковица – типичный орган вегетативного возобновления и размножения. Луковицы наиболее характерны для растений из семейств лилейных (лилии, тюльпаны), луковых (луки) и амариллисовых (нарциссы, гиацинты).
Клубнелуковица внешне напоминает луковицу, но ее чешуевидные листья не являются запасающими; они сухие и пленчатые, а запасные вещества откладываются в утолщенной стеблевой части (шафран, гладиолус).

Рис. 4.24. Подземные метаморфозы побега: 1, 2, 3, 4 – последовательность развития и строение клубня картофеля; 5 – клубень цикламена; 6 – клубень кольраби; 7 – луковички лилии тигровой; 8 – луковица лука репчатого; 9 – луковица лилии; 10 – участок длинного корневища пырея ползучего.
Видоизменяться могут не только подземные, но и надземные побеги растений (рис. 4.25). Довольно часто встречаются надземные столоны. Это плагиотропные недолговечные побеги, функция которых вегетативное размножение, расселение и захват территории. Если столоны несут зеленые листья и участвуют в процессе фотосинтеза, их называют плетями (костяника, живучка ползучая). У земляники столоны лишены развитых зеленых листьев, стебли у них тонкие и хрупкие, с очень длинными междоузлиями. Такие более высокоспециализированные для функции вегетативного размножения столоны называют усами.
Сочными, мясистыми, приспособленными для накопления воды могут быть не только луковицы, но и надземные побеги, обычно у растений, живущих в условиях недостатка влаги. Водозапасающими органами могут быть листья или стебли, иногда даже почки. Такие сочные растения носят название суккулентов. Листовые суккуленты запасают воду в тканях листьев (алоэ, агавы, толстянки, родиола, или золотой корень). Стеблевые суккуленты характерны для американского семейства кактусовых и африканских молочайных. Сочный стебель выполняет водозапасающую и ассимилирующую функцию; листья редуцированы или превращены в колючки (рис. 4.25, 1). У большинства кактусов стебли колонновидные или шаровидные, листьев на них совсем не образуется, но узлы хорошо заметны по расположению пазушных побегов – ареол, имеющих вид бородавок или удлиненных выростов с колючками или пучками волосков. Превращение листьев в колючки уменьшает испаряющую поверхность растения и защищает от поедания животными. Примером метаморфоза почки в суккулентный орган – кочан служит культурная капуста.
Колючки кактусов имеют листовое происхождение. Листовые колючки нередко встречаются и у несуккулентных растений (барбарис) (рис. 4.26, 1). У многих растений колючки имеют не листовое, а стеблевое происхождение. У дикой яблони, дикой груши, жостера слабительного в колючки метаморфизируются укороченные побеги, имеющие ограниченный рост и оканчивающиеся острием. Вид жесткой одревесневшей колючки они приобретают после опадения листьев. У боярышника (рис. 4.26, 3) колючки, образующиеся в пазухах листьев, совершенно безлистны с самого начала. У гледичии (рис. 4.25, 5) мощные разветвленные колючки образуются на стволах из спящих почек. Образование колючек любого происхождения, как правило, есть результат недостатка влаги. При выращивании многих колючих растений в искусственной влажной атмосфере они утрачивают колючки: вместо них вырастают нормальные листья (верблюжья колючка) или облиственные побеги (английский дрок).
Побеги ряда растений несут шипы. Шипы отличаются от колючек меньшими размерами, это выросты – эмергенцы – покровной ткани и тканей коры стебля (шиповник, крыжовник) (рис. 4.26, 4).
Приспособление к недостатку влаги очень часто выражается в ранней утрате, метаморфозе или редукции листьев, теряющих основную функцию фотосинтеза. Это компенсируется тем, что роль ассимилирующего органа берет на себя стебель. Иногда такой ассимилирующий стебель безлистного побега остается внешне неизмененным (испанский дрок, верблюжья колючка) (рис. 4.25, 3). Дальнейшим шагом в указанной смене функций является формирование таких органов, как филлокладии и кладодии. Это уплощенные листоподобные стебли или целые побеги. На побегах иглицы (рис. 4.25, 4), в пазухах чешуевидных листьев, развиваются плоские листовидные филлокладии, имеющие, подобно листу, ограниченный рост. На филлокладиях образуются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях, значит, филлокладий соответствует целому пазушному побегу. Мелкие, игольчатые филлокладии образуются у спаржи в пазухах чешуевидных листьев основного скелетного побега. Кладодии – это уплощенные стебли, сохраняющие, в отличие от филлокладиев, способность к длительному росту.
Для некоторых растений характерно видоизменение листьев или их частей, а иногда целых побегов в усики, которые закручиваются вокруг опоры, помогая тонкому и слабому стеблю сохранять вертикальное положение. У многих бобовых в усики превращается верхняя часть перистосложного листа (горох, горошки, чина). В других случаях в усики превращаются прилистники (сассапариль). Очень характерные усики листового происхождения формируются у тыквенных, причем можно видеть все переходы от нормальных листьев к полностью метаморфизированным. Усики побегового происхождения можно наблюдать у винограда (рис. 4.25, 2), пассифлоры и ряда других растений.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 1889; Нарушение авторских прав?; Мы поможем в написании вашей работы!