КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вегетативные органы
|
|
|
|
Вегетативные органы растений — части растения, выполняющие основные функции питания и обмена веществ с внешней средой. Вегетативные органы бывают растущими и выросшими. Именно поэтому они часто находятся на разных стадиях развития, хотя принадлежат одному растению.
К вегетативным органам относятся:
- листостебельные побеги, обеспечивающие фотосинтез;
- корни, обеспечивающие водоснабжение и минеральное питание.
Вегетативные органы могут выполнять функции вегетативного размножения.
| Клеточная оболочка растительной клетки |
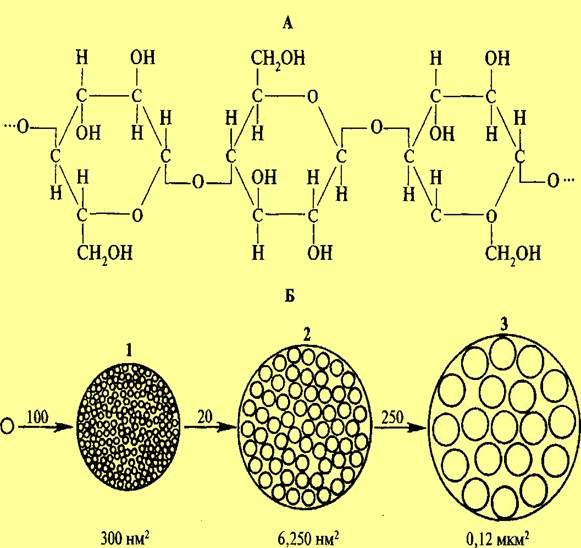
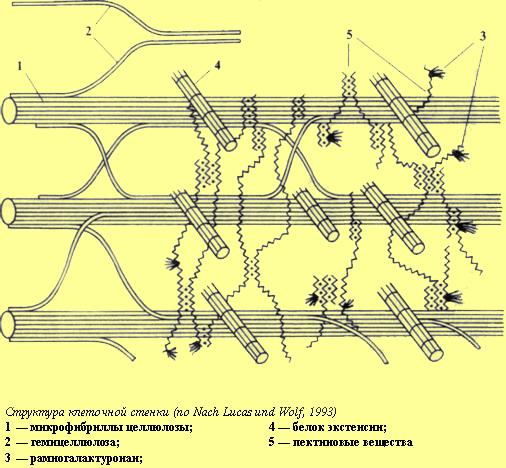
Характерной особенностью растительной клетки является наличие жесткой (твердой) клеточной стенки. Клеточная оболочка определяет форму клетки, придает клеткам и тканям растений механическую прочность и опору, защищает цитоплазматическую мембрану от разрушения под влиянием гидростатического давления, развиваемого внутри клетки. Однако такую оболочку нельзя рассматривать только как механический каркас. Клеточная оболочка обладает такими свойствами, которые позволяют противостоять давлению воды внутри клетки, и в то же время обладает растяжимостью и способностью к росту. Она является противоинфекционным барьером, принимает участие в поглощении минеральных веществ, являясь своеобразным ионообменником. Появились данные, что углеводные компоненты клеточной оболочки, взаимодействуя с гормонами, вызывают ряд физиологических изменений. Для молодых растущих клеток характерна первичная клеточная оболочка. По мере их старения образуется вторичная структура. Первичная клеточная оболочка, как правило, малоспециализирована, имеет более простое строение и меньшую толщину, чем вторичная. В состав клеточной оболочки входят целлюлоза, гемицеллюлозы, пектиновые вещества, липиды и небольшое количество белка. Компоненты клеточной оболочки являются продуктами жизнедеятельности клетки. Они выделяются из цитоплазмы и претерпевают превращения на поверхности плазмалеммы. Первичные клеточные стенки содержат из расчета на сухое вещество: 25% целлюлозы, 25% гемицеллюлозы, 35% пектиновых веществ и 1—8% структурных белков. Однако цифры весьма колеблются. Так, в состав клеточных стенок колеоптилей злаков входит до 60—70% гемицеллюлоз, 20—25 % целлюлозы, 10% пектиновых веществ. Вместе с тем клеточные стенки эндосперма содержат до 85% гемицеллюлоз. Во вторичных клеточных стенках больше целлюлозы. Остов клеточной оболочки составляют переплетенные микро- и макрофибриллы целлюлозы. Целлюлоза, или клетчатка (С6Н10О5)n, представляет собой длинные неразветвленные цепочки, состоящие из 3—10 тыс. остатков D-глюкозы, соединенных b -1,4-гликозидными связями. Молекулы целлюлозы объединены в мицеллу, мицеллы объединены в микрофибриллу, микрофибриллы объединены в макрофибриллу. Макрофибриллы, мицеллы и микрофибриллы соединены в пучки водородными связями. Диаметр мицеллы составляет 5 нм, диаметр микрофибриллы — 25—30 нм, макрофибриллы — 0,5 мкм. Структура микро- и макрофибрилл неоднородна. Наряду с хорошо организованными кристаллическими участками имеются паракристаллические, аморфные.
Микро- и макрофибриллы целлюлозы в клеточной оболочке погружены в аморфную желеобразную массу — матрикс. Матрикс состоит из гемицеллюлоз, пектиновых веществ и белка. Гемицеллюлозы, или полуклетчатки,— это производные пентоз и гексоз. Степень полимеризации у этих соединений меньше по сравнению с клетчаткой (150—300 мономеров, соединенные b -1,3- и b -1,4-гли-козидными связями). Из гемицеллюлоз наибольшее значение имеют ксило-глюканы, которые входят в состав матрикса первичной клеточной стенки. Это цепочки остатков D-глюкозы, соединенных b -1,4-гликозидными связями, у которых от шестого углеродного атома глюкозы отходят боковые цепи, главным образом из остатков D-ксилозы. К ксилозе могут присоединяться остатки галактозы и фукозы. Гемицеллюлозы способны связываться с целлюлозой, поэтому они формируют вокруг микрофибрилл целлюлозы оболочку, скрепляя их в сложную цепь.
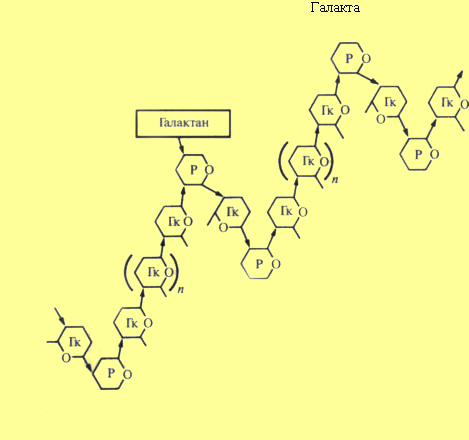
 Пектиновые вещества — это полимерные соединения углеводного типа. Они обусловливают высокую оводненность клеточной оболочки. Важнейшим представителем пектиновых веществ являются рамногалактуронаны, представляющие собой цепочку остатков oc-D-галактуроновой кислоты (Гк), к которой в ряде мест присоединяются остатки рамнозы. Рамноза (С6Н1205) — производное глюкозы. Вследствие внедрения рамнозы основная цепь этого полисахарида приобретает зигзагообразную форму. В некоторых случаях четвертый углеродный атом рамнозы замещен на галактозу. Пектиновые вещества содержат большое количество карбоксильных групп и могут эффективно связывать ионы двухвалентных металлов, например, Са+2, что играет роль в объединении компонентов клеточной стенки. Ионы Са+2 могут обмениваться на такие ионы как К+ и Н+, что обеспечивает катионообменную способность.
Клеточные стенки содержат также белок экстенсии (до 10%). Это гликопро-теид, у которого около 30% всех аминокислот белковой части предстаачено оксипролином. К оксипролину присоединяются углеводные цепочки, состоящие из четырех остатков моносахара арабинозы. По исследованиям Д. Лампорта, именно цепочки арабинозы придают устойчивость структуре экстенсина. Вместе с тем экстенсии является связующим звеном между полисахаридами, входящими в состав клеточной оболочки, соединяя их в единый каркас. Наряду с этим в состав клеточной оболочки входят специфические углевод-связывающие белки пектины, согласно современным представлениям участвующие в обеспечении узнавания и взаимодействия клеток, рецепторных свойств, защиты от инфекций. В клеточных оболочках локализован ряд ферментов, по преимуществу гидролаз (глюкозидазы, гликозидазы и др.). Эти ферменты, расщепляя соответствующие связи, могут участвовать в растяжении клеточной оболочки.
Пектиновые вещества — это полимерные соединения углеводного типа. Они обусловливают высокую оводненность клеточной оболочки. Важнейшим представителем пектиновых веществ являются рамногалактуронаны, представляющие собой цепочку остатков oc-D-галактуроновой кислоты (Гк), к которой в ряде мест присоединяются остатки рамнозы. Рамноза (С6Н1205) — производное глюкозы. Вследствие внедрения рамнозы основная цепь этого полисахарида приобретает зигзагообразную форму. В некоторых случаях четвертый углеродный атом рамнозы замещен на галактозу. Пектиновые вещества содержат большое количество карбоксильных групп и могут эффективно связывать ионы двухвалентных металлов, например, Са+2, что играет роль в объединении компонентов клеточной стенки. Ионы Са+2 могут обмениваться на такие ионы как К+ и Н+, что обеспечивает катионообменную способность.
Клеточные стенки содержат также белок экстенсии (до 10%). Это гликопро-теид, у которого около 30% всех аминокислот белковой части предстаачено оксипролином. К оксипролину присоединяются углеводные цепочки, состоящие из четырех остатков моносахара арабинозы. По исследованиям Д. Лампорта, именно цепочки арабинозы придают устойчивость структуре экстенсина. Вместе с тем экстенсии является связующим звеном между полисахаридами, входящими в состав клеточной оболочки, соединяя их в единый каркас. Наряду с этим в состав клеточной оболочки входят специфические углевод-связывающие белки пектины, согласно современным представлениям участвующие в обеспечении узнавания и взаимодействия клеток, рецепторных свойств, защиты от инфекций. В клеточных оболочках локализован ряд ферментов, по преимуществу гидролаз (глюкозидазы, гликозидазы и др.). Эти ферменты, расщепляя соответствующие связи, могут участвовать в растяжении клеточной оболочки.
 Клеточная оболочка способна к утолщению и видоизменению. В результате этого образуется ее вторичная структура. Утолщение оболочки происходит путем наложения новых слоев на первичную оболочку. Ввиду того, что наложение идет уже на твердую оболочку, фибриллы целлюлозы в каждом слое лежат параллельно, а в соседних слоях — под углом друг к другу. Предполагается, что за ориентацию микрофибрилл целлюлозы ответственны микротрубочки. Этим достигается значительная прочность (и твердость) вторичной оболочки. По мере того как число слоев фибрилл целлюлозы становится больше, и толщина стенки увеличивается, она теряет эластичность и способность к росту. Во вторичной клеточной стенке содержание целлюлозы значительно возрастает (в некоторых случаях до 60% и более). По мере дальнейшего старения клеток матрикс оболочки может заполняться различными веществами — лигнином, суберином. Лигнин — это полимер, образующийся путем конденсации ароматических спиртов. Включение лигнина сопровождается одревеснением, увеличением прочности и уменьшением растяжимости. Мономерами суберина являются насыщенные и ненасыщенные оксожирные кислоты. Пропитанные суберином клеточные стенки (опробковение оболочки) становятся труднопроницаемыми для воды и растворов. На поверхности клеточной стенки могут откладываться кутин и воск. Кутин состоит из оксожирных кислот и их солей, выделяется через клеточную стенку на поверхность эпидермальной клетки и участвует в образовании кутикулы. В состав кутикулы могут входить воска, которые также секретирует цитоплазма. Кутикула препятствует испарению воды, регулирует водно-тепловой режим тканей растений.
Исследования позволили дать предположительную модель взаимосвязи и взаиморасположения всех перечисленных веществ в клеточной стенке. Согласно этой модели в первичной клеточной оболочке микрофибриллы целлюлозы располагаются либо беспорядочно, либо перпендикулярно (в основном) продольной оси клетки. Между микрофибриллами целлюлозы находятся молекулы гемицеллюлозы, которые, в свою очередь, связаны через пектиновые вещества с белком. При этом последовательность веществ следующая: целлюлоза — гемицеллюлозы — пектиновые вещества — белок — пектиновые вещества — гемицеллюлозы — целлюлоза. Микрофибриллы целлюлозы и вещества матрикса оболочки связаны между собой. Единственными нековалентными связями являются водородные между целлюлозными микрофибриллами и гемицеллюлозой (по преимуществу ксилоглюканом). Между ксилоглюканом и пектиновыми веществами, так же как и между пектиновыми веществами и белком экстенсином, возникают ковалентные связи.
Клеточная оболочка способна к утолщению и видоизменению. В результате этого образуется ее вторичная структура. Утолщение оболочки происходит путем наложения новых слоев на первичную оболочку. Ввиду того, что наложение идет уже на твердую оболочку, фибриллы целлюлозы в каждом слое лежат параллельно, а в соседних слоях — под углом друг к другу. Предполагается, что за ориентацию микрофибрилл целлюлозы ответственны микротрубочки. Этим достигается значительная прочность (и твердость) вторичной оболочки. По мере того как число слоев фибрилл целлюлозы становится больше, и толщина стенки увеличивается, она теряет эластичность и способность к росту. Во вторичной клеточной стенке содержание целлюлозы значительно возрастает (в некоторых случаях до 60% и более). По мере дальнейшего старения клеток матрикс оболочки может заполняться различными веществами — лигнином, суберином. Лигнин — это полимер, образующийся путем конденсации ароматических спиртов. Включение лигнина сопровождается одревеснением, увеличением прочности и уменьшением растяжимости. Мономерами суберина являются насыщенные и ненасыщенные оксожирные кислоты. Пропитанные суберином клеточные стенки (опробковение оболочки) становятся труднопроницаемыми для воды и растворов. На поверхности клеточной стенки могут откладываться кутин и воск. Кутин состоит из оксожирных кислот и их солей, выделяется через клеточную стенку на поверхность эпидермальной клетки и участвует в образовании кутикулы. В состав кутикулы могут входить воска, которые также секретирует цитоплазма. Кутикула препятствует испарению воды, регулирует водно-тепловой режим тканей растений.
Исследования позволили дать предположительную модель взаимосвязи и взаиморасположения всех перечисленных веществ в клеточной стенке. Согласно этой модели в первичной клеточной оболочке микрофибриллы целлюлозы располагаются либо беспорядочно, либо перпендикулярно (в основном) продольной оси клетки. Между микрофибриллами целлюлозы находятся молекулы гемицеллюлозы, которые, в свою очередь, связаны через пектиновые вещества с белком. При этом последовательность веществ следующая: целлюлоза — гемицеллюлозы — пектиновые вещества — белок — пектиновые вещества — гемицеллюлозы — целлюлоза. Микрофибриллы целлюлозы и вещества матрикса оболочки связаны между собой. Единственными нековалентными связями являются водородные между целлюлозными микрофибриллами и гемицеллюлозой (по преимуществу ксилоглюканом). Между ксилоглюканом и пектиновыми веществами, так же как и между пектиновыми веществами и белком экстенсином, возникают ковалентные связи.
 Клеточная стенка растительной клетки пронизана плазмодесмами. В клеточной стенке они могут располагаться равномерно или группами. Плазмодесмы обнаружены в клетках всех групп растений, за исключением репродуктивных клеток. На каждые 100 мкм2 клеточной оболочки имеется примерно 10—30 плазмодесм. Плазмодесма представляет собой канал (пору) шириной до 1 мкм, выстланный плазмалеммой. В центре поры имеется десмотрубка, которая образована мембранами эндоплазматической сети соседних клеток. Десмотрубка окружена белками и слоем цитоплазмы, которая соединяется с цитоплазмами соседних клеток. Благодаря плазмодесмам цитоплазма всех объединена в единое целое — симпласт. Взаимосвязанная система клеточных стенок и межклеточных промежутков называется апопласт (свободное пространство). Симпласт и апопласт являются важнейшими путями передвижения воды и минеральных веществ между клетками. Одним из путей регуляции транспортной функции является подвижность структуры плазмодесм (Ю.В. Гамалей) и объем свободного пространства. Толщина клеточной стенки колеблется у разных видов растений от десятых долей до 10 мкм. Так, клетки кортикальной паренхимы более тонкие, а специализированные клетки эпидермиса, ксилемы, флоэмы и другие — более толстые. У клетки отдельные стороны клеточной стенки могут различаться по толщине, количеству плазмодесм. Клеточная стенка внешней стороны клетки эпидермиса толще, имеет меньше плазмодесм, чем внутренняя сторона этой клетки.
Между клеточными оболочками двух соседних клеток в местах их соприкосновения имеется так называемая срединная пластинка; в состав срединной пластинки входят пектиновые вещества, главным образом в виде пектатов кальция (кальциевая соль пектиновой кислоты). Эти вещества как бы цементируют, склеивают растущие клетки. При недостатке кальция пектиновые вещества превращаются в слизь, наблюдается ослизнение ткани, и клетки разъединяются (мацерация ткани). При созревании плодов пектиновые вещества срединных пластинок, склеивающие клетки, переходят в растворимую форму и благодаря этому плоды становятся мягкими. Срединная пластинка является первым слоем, образующимся при делении клетки. Клеточная оболочка способна к эластическому (обратимому) и пластическому (необратимому) растяжению. Эластическое растяжение происходит под влиянием развивающегося в клетке давления воды (тургорного давления). Макрофибриллы целлюлозы не связаны между собой и скреплены только матриксом. Они под влиянием давления как бы раздвигаются, клеточная оболочка становится тоньше. Пластическое необратимое растяжение это собственно рост клеточной оболочки. Рост клеточной оболочки начинается с ее разрыхления. Поскольку микрофибриллы практически не растягиваются в длину, то для того, чтобы произошло растяжение оболочки, они должны скользить вдоль оси растяжения, удаляясь друг от друга. Способность микрофибрилл скользить друг около друга очень важна для обеспечения роста растяжением. Чем это скольжение проходит легче, тем пластичнее клеточная стенка. Легкость скольжения обусловлена водородными связями между микрофибриллами целлюлозы и ксилоглюканом (пектиновыми веществами). Особенностью этих связей является их лабильность — легкое разрушение и возобновление, не требующее значительных энергетических затрат. После того как растяжение клетки произошло, между вновь образовавшимися микрофибриллами целлюлозы и веществами матрикса возникают связи. Подкисление увеличивает растяжимость клеток. В этом важную роль играет экстенсии. Показано, что этот белок катализирует зависимое от рН растяжение клеточных стенок. На размягчение клеточных стенок также влияют ферменты глюканазы, которые вызывают распад ксилоглюкана. Клеточные стенки неактивны и достаточно устойчивы, что и обеспечивает выполнение механической и защитной функции. Однако под влиянием патогенов они могут в течение секунд модифицироваться. Причем, молекулы, образующиеся при распаде материала клеточных стенок, играют роль сигналов, информируя растительный организм об инфекции и выполняя защитную роль. Показано, что разрушение клеточных стенок может приводить к накоплению олигосахаридов, способных стимулировать синтез сигнальных молекул (фито-алексинов, этилена и др.), которые в соединении с патогеном обусловливают дальнейший сигнал, рост и морфогенез у изолированных частей (И.А. Тарчевский).
Клеточная стенка растительной клетки пронизана плазмодесмами. В клеточной стенке они могут располагаться равномерно или группами. Плазмодесмы обнаружены в клетках всех групп растений, за исключением репродуктивных клеток. На каждые 100 мкм2 клеточной оболочки имеется примерно 10—30 плазмодесм. Плазмодесма представляет собой канал (пору) шириной до 1 мкм, выстланный плазмалеммой. В центре поры имеется десмотрубка, которая образована мембранами эндоплазматической сети соседних клеток. Десмотрубка окружена белками и слоем цитоплазмы, которая соединяется с цитоплазмами соседних клеток. Благодаря плазмодесмам цитоплазма всех объединена в единое целое — симпласт. Взаимосвязанная система клеточных стенок и межклеточных промежутков называется апопласт (свободное пространство). Симпласт и апопласт являются важнейшими путями передвижения воды и минеральных веществ между клетками. Одним из путей регуляции транспортной функции является подвижность структуры плазмодесм (Ю.В. Гамалей) и объем свободного пространства. Толщина клеточной стенки колеблется у разных видов растений от десятых долей до 10 мкм. Так, клетки кортикальной паренхимы более тонкие, а специализированные клетки эпидермиса, ксилемы, флоэмы и другие — более толстые. У клетки отдельные стороны клеточной стенки могут различаться по толщине, количеству плазмодесм. Клеточная стенка внешней стороны клетки эпидермиса толще, имеет меньше плазмодесм, чем внутренняя сторона этой клетки.
Между клеточными оболочками двух соседних клеток в местах их соприкосновения имеется так называемая срединная пластинка; в состав срединной пластинки входят пектиновые вещества, главным образом в виде пектатов кальция (кальциевая соль пектиновой кислоты). Эти вещества как бы цементируют, склеивают растущие клетки. При недостатке кальция пектиновые вещества превращаются в слизь, наблюдается ослизнение ткани, и клетки разъединяются (мацерация ткани). При созревании плодов пектиновые вещества срединных пластинок, склеивающие клетки, переходят в растворимую форму и благодаря этому плоды становятся мягкими. Срединная пластинка является первым слоем, образующимся при делении клетки. Клеточная оболочка способна к эластическому (обратимому) и пластическому (необратимому) растяжению. Эластическое растяжение происходит под влиянием развивающегося в клетке давления воды (тургорного давления). Макрофибриллы целлюлозы не связаны между собой и скреплены только матриксом. Они под влиянием давления как бы раздвигаются, клеточная оболочка становится тоньше. Пластическое необратимое растяжение это собственно рост клеточной оболочки. Рост клеточной оболочки начинается с ее разрыхления. Поскольку микрофибриллы практически не растягиваются в длину, то для того, чтобы произошло растяжение оболочки, они должны скользить вдоль оси растяжения, удаляясь друг от друга. Способность микрофибрилл скользить друг около друга очень важна для обеспечения роста растяжением. Чем это скольжение проходит легче, тем пластичнее клеточная стенка. Легкость скольжения обусловлена водородными связями между микрофибриллами целлюлозы и ксилоглюканом (пектиновыми веществами). Особенностью этих связей является их лабильность — легкое разрушение и возобновление, не требующее значительных энергетических затрат. После того как растяжение клетки произошло, между вновь образовавшимися микрофибриллами целлюлозы и веществами матрикса возникают связи. Подкисление увеличивает растяжимость клеток. В этом важную роль играет экстенсии. Показано, что этот белок катализирует зависимое от рН растяжение клеточных стенок. На размягчение клеточных стенок также влияют ферменты глюканазы, которые вызывают распад ксилоглюкана. Клеточные стенки неактивны и достаточно устойчивы, что и обеспечивает выполнение механической и защитной функции. Однако под влиянием патогенов они могут в течение секунд модифицироваться. Причем, молекулы, образующиеся при распаде материала клеточных стенок, играют роль сигналов, информируя растительный организм об инфекции и выполняя защитную роль. Показано, что разрушение клеточных стенок может приводить к накоплению олигосахаридов, способных стимулировать синтез сигнальных молекул (фито-алексинов, этилена и др.), которые в соединении с патогеном обусловливают дальнейший сигнал, рост и морфогенез у изолированных частей (И.А. Тарчевский).
|
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 593; Нарушение авторских прав?; Мы поможем в написании вашей работы!