КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Гормоны
|
|
|
|
Вопрос № 3.
Гормоны – органические сигнальные молекулы беспроводного системного действия. Гормоны – органические вещества, синтезирующиеся в эндокринных железах, транспортируемые кровью и действующие на ткани мишени (гормоны щитовидной железы, надпочечников, поджелудочной железы и т.д). Всего известно более 100 гормонов.
Гормоноподобные вещества - органические вещества, синтезирующиеся апудоцитами, транспортируемые кровью и действующие на ткани мишени. Апудоциты – это диффузные эндокриноциты (отдельные клетки, не оформленные в железу), они образуются из эктодермы, эндодермы или мезодермы. Апудоциты формируют АПУД систему (диффузную гормональную систему). Апудоциты находятся в ЖКТ, вилочковой железе, сердце, печени, почках, ЦНС, плаценте и коже. Часто гормоноподобные вещества имеют то же самое строение, что и истинные гормоны, нейромедиаторы. Например, в ЖКТ синтезируются вазоактивный интестинальный пептид (ВИП), холецистокинин, гастрин, нейротензин, мет-, лейэнкефалин и др..
Тканевые гормоны - органические вещества, синтезирующиеся отдельными клетками, не транспортируемые кровью и действующие на ткани мишени.
Особенности действия гормонов:
1. Действуют в малых количествах (10-6-10-12 ммоль/л);
2. Существует абсолютная или высокая специфичность в действии гормонов.
3. Переносят только информацию. Не используются в энергетических и строительных целях;
4. Действуют опосредованно через рецепторы и внутриклеточные посредники (Са2+, цАМФ, цГМФ, ДАГ, ИФ3 и т.д.). Например, через аденилатциклазную, инозитолтрифосфатную системы;
5. Регулируют активность или количество ферментов;
6. Зависят от ЦНС;
7. Беспороговый принцип. Даже 1 молекула гормона способна оказать эффект;
8. Пермессивность действия. Конечный эффект - результат действия множества гормонов.
Гормоны могут оказывать как системное, так и местное действие.
Эндокринное (системное) действие гормонов (эндокринный эффект) реализуется, когда они транспортируются кровью и контактируют с органами и тканями всего организма. Характерно для истинных гормонов и гормоноподобных веществ.
Местное действие гормонов реализуется, когда они влияют на клетки, в которых были синтезированы (аутокринный эффект), или на соседние клетки (паракринный эффект). Характерно для тканевых гормонов, есть также у истинных гормонов и гормоноподобных веществ.
I. Обмен белковых (пептидных) гормонов
1. Синтез гормонов происходит на рибосомах.
2. Активация. Гормоны синтезируются, как правило, в неактивной форме. Активация происходит в ЭПС, аппарате Гольджи, секреторных гранулах или на периферии (в крови).
3. Хранение. Гормоны накапливаются и хранятся в секреторных гранулах. Запас инсулина на 5 дней, у других гормонов как правило меньше.
4. Секреция гормонов происходит при активном экзоцитозе секреторных гранул. Секрецию стимулируют нейромедиаторы, метаболиты и другие гормоны.
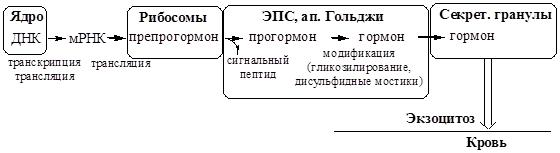
5. Транспорт белковых гормонов осуществляется в основном в свободном виде, т.к. они водорастворимы. Часть гормонов транспортируется в комплексе с белками.
6. Действие гормонов. Белковые гормоны взаимодействуют с мембранными рецепторами и через систему внутриклеточных посредников регулируют активность ферментов, что влияет на интенсивность метаболизма в тканях мишенях. Механизм действия гормонов (рецепторы, посредники) детально рассмотрен в разделе ферменты.
7. Инактивация. Гормоны инактивируются гидролизом до АК в тканях мишенях, печени, почках и т.д. Время полураспада инсулина, глюкагона ТЅ = 3-5мин, у СТГ ТЅ= 50 мин.
Детально обмен белковых гормонов рассмотрен в теме «углеводы» на примере инсулина.
II. Обмен стероидных гормонов
1. Синтез гормонов происходит из холестерина в гладком ЭПР и митохондриях коры надпочечников, гонадах, коже, печени, почках. Превращение стероидов состоит в отщеплении алифатической боковой цепи, гидроксилировании, дегидрировании, изомеризации, либо в ароматизации кольца.
2. Активация. Стероидные гормоны часто образуются уже в активном виде.
3. Хранение. Синтезированные гормоны накапливаются в цитоплазме в комплексе со специальными белками.
4. Секреция стероидных гормонов происходит пассивно. Гормоны переходят с цитоплазматических белков в клеточную мембрану, откуда их забирают транспортные белки крови.
5. Транспорт. Стероидные гормоны, т.к. они водонерастворимы, переносятся в крови преимущественно в комплексе с транспортными белками (альбумины).
6. Действие гормонов. Стероидные гормоны взаимодействуют с цитоплазматическими и ядерными рецепторами и регулируют количество ферментов, что влияет на интенсивность метаболизма в тканях мишенях. Механизм действия гормонов детально рассмотрен в разделе ферменты.
7. Инактивация. Стероидные гормоны инактивируются так же как и ксенобиотики реакциями гидроксилирования и конъюгации в печени и тканях мишенях. Инактивированные производные выводятся из организма с мочой и желчью. Период полураспада в крови обычно больше пептидных гормонов. У кортизола ТЅ = 1,5-2 часа.
Механизмы передачи гормональных сигналов в клетки
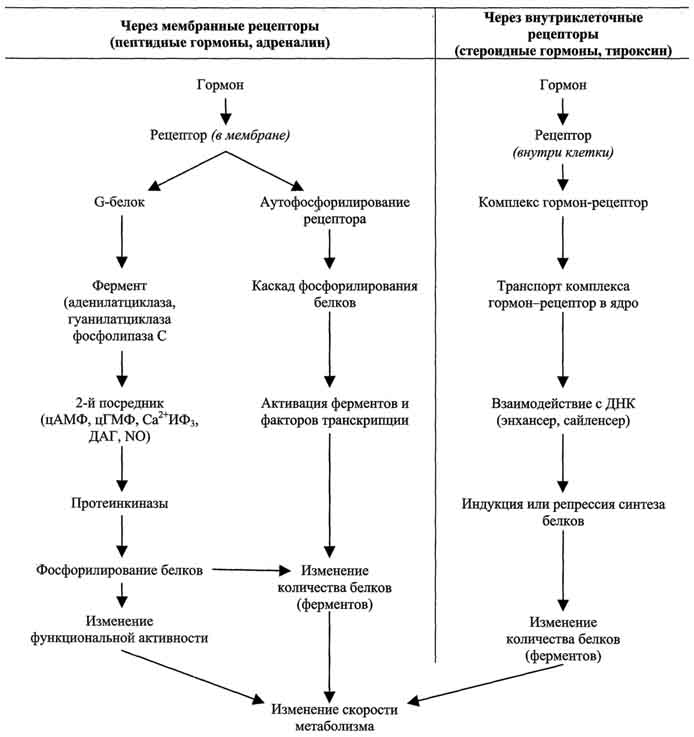
По механизму действия гормоны можно разделить на 2 группы. К первой группе относят гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды). Вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами.
Связывание гормона (первичного посредника) с рецептором приводит к изменению кон-формации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма:
в результате изменения активности ферментов;
в результате изменения количества ферментов.
1. Передача гормональных сигналов
через мембранные рецепторы
Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрацииособых молекул внутри клетки - вторичных посредников. Вторичными посредниками могут быть следующие молекулы: цАМФ, цГМФ, ИФ3, ДАТ, Са2+, NO.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ, действуют через трёхкомпонентную систему, которая включает белок-рецептор, G-белок и фермент аденилатциклазу. Образующийся под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилируюшую ферменты и другие белки. Известно более 200 различных G-белков, в структуре которых обнаружены 3 субъединицы α, β и γ . В отсутствие гормона α-субъединица G-белка связана с ГДФ. Образование комплекса гормонрецептора приводит к конформационным изменениям α-субъединицы, замене ГДФ на ГТФ и отщеплению димера βγ от α-ГТФ. В случае рецепторов, сопряжённых с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу.
В случае рецепторов, сопряжённых с Giбелком, субъединица αi-ГТФ ингибирует аденилатциклазу.
Другая система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Цитоплазматический домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ> (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-за-висимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. Например, фосфодиэстерааа, которая гидролизует цАМФ до АМФ, активируется в результате фосфорилирования цГМФ-зависимой протеинкиназой.
Некоторые гормоны (например, вазопрессин или адреналин), образуя комплекс с соответствующими рецепторами (рецептор V1 для вазопрессина и αi-рецептор для адреналина), через активацию соответствующих G-белков активируют фосфолипазу С, в результате чего в клетке появляются вторичные посредники ИФ3, ДАГ. Молекула ИФ3 стимулирует высвобождение Са2+ из ЭР. Кальций связывается с белком кальмодулином. Этот комплекс активирует Са2+-кальмодулинзависимую протеинкиназу. Ионы кальция и ДАГ участвуют в активации протеинкиназы С.
Многие гормоны передают сигнал в клетку через рецепторы, которые либо обладают тиро-зинкиназной активностью, либо связываются с цитоплазматическими белками, проявляющими активность тирозинкиназы. Связывание инсулина с мембранным рецептором, который является тирозинкиназой и имеет центр фосфорилирования, инициирует аутофосфорилирование и последующее фосфорилирование субстратов рецептора инсулина и других белков.
В случае взаимодействия, например, эпидермального фактора роста или инсулиноподобного фактора роста -1с мембранным рецептором сначала происходят димеризация рецептора и его активация. Активированный таким образом гомодимер рецептора, участок которого на внутренней стороне мембраны обладает активностью тирозинкиназы, фосфорилируется сам (аутофосфорилирование) и вызывает фосфорилирование других белков и ферментов, которые участвуют в активации факторов транскрипции генов.
Некоторые гормоны (например, гормон роста, пролактин, интерферон, цитокины) взаимодействуют с мембранными рецепторами, ассоциированными с цитоплазматическими протеинкиназами (так называемыми "Янус-киназами", или киназами семейства JAK). Присоединение гормона вызьшает димеризацию рецептора, присоединение Янус-киназ, их аутофосфорилирование и активацию. Янус-киназы, в свою очередь, фос-форилируют рецептор по остаткам тирозина, в результате чего рецептор связывается с другими белками, например, особыми белками - переносчиками сигнала и активаторами транскрипции (ПСАТ, или STAT - от англ, signal transducer and activator of transcription - переносчик сигнала и активатор транскрипции). Далее следует инициируемый тирозинкиназой каскад реакций фосфорилирования. Белки STAT фосфорилируются, образуют димеры, транспортируются в ядро, где, связываясь со специфическими участками ДНК, участвуют в регуляции транскрипции.
Сигнальной молекулой в клетке может служить также оксид азота NO, образующийся в организме из аргинина при участии фермента NO-синтазы, присутствующего в нервной ткани, эндотелии сосудов, тромбоцитах и других тканях. Молекула NO может быстро диффундировать через мембрану эндотелиальных клеток, где она синтезируется, в соседние клетки. Действие оксида азота кратковременно, так как Т1/2 NO колеблется в пределах 5-10 с. В крови молекула существует примерно 100 мс, поскольку быстро взаимодействует с молекулярным кислородом, образуя нитрит, который далее превращается в нитрат и экскретируется с мочой. В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в активный центр гуанилатциклазы ионом железа, способствуя тем самым быстрому образованию цГМФ. Увеличение концентрации цГМФ в клетках гладких мышц вызывает активацию киназ, что в конечном итоге приводит к расслаблению ГМК сосудов и последующему их расширению. Механизм действия оксида азота объясняет использование нитроглицерина в качестве лекарственного препарата для снятия острых болей в сердце, поскольку нитроглицерин - источник образующихся молекул NO, которые и вызывают расслабление кровеносных сосудов и увеличение притока крови в миокард.
2. Передача сигналов через внутриклеточные
рецепторы
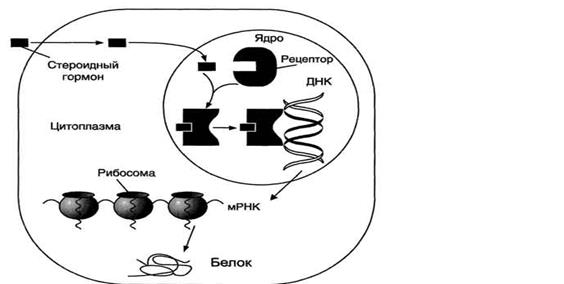
Стероидные и тиреоидные гормоны связываются с рецепторами внутри клетки и регулируют скорость транскрипции специфических генов (рис. 11-5).
В отсутствие гормона внутриклеточные рецепторы связаны обычно с другими белками в цитозоле или ядре. Например, рецепторы глюкокортикоидов образуют в цитозоле комплекс с шапероном, что препятствует связыванию рецептора с молекулой ДНК.
Взаимодействие гормона с центром связывания на С-концевом участке полипептидной цепи рецептора вызывает конформационные изменения и освобождение рецептора от шаперона. Происходит объединение 2 молекул рецептора с образованием гомодимера. Димер рецептора узнаёт специфическую последовательность нуклеотидов, которая расположена в промоторной области гена. Взаимодействие со специфическим участком ДНК HRE (от англ, hormone response element, элемент, реагирующий на воздействие гормона) обеспечивает центральный домен рецептора. Этот домен содержит аминокислотную последовательность, образующую 2 "цинковых пальца". В каждом "цинковом пальце" атом цинка связан с 4 остатками цистеина.
В структуре одного "цинкового пальца" имеется последовательность аминокислот, отвечающая за связывание с ДНК, а второй "цинковый палец" содержит последовательность аминокислот, участвующую в димеризации рецепторов. Взаимодействие комплекса гормон-рецептор с определённой последовательностью нуклеотидрв в промоторной части ДНК приводит к активации транскрипции.
Рецепторы тиреоидных гормонов всегда связаны с ДНК. В отсутствие гормонов соответствующие рецепторы ингибируют экспрессию генов. Напротив, взаимодействие с гормоном превращает их в активаторы транскрипции.
Рис. 11-5. Передача гормональных сигналов через внутриклеточные рецепторы (рецепторы стероидных гормонов могут находиться в цитоплазме и ядре).
.
|
|
|
|
|
Дата добавления: 2015-05-08; Просмотров: 1124; Нарушение авторских прав?; Мы поможем в написании вашей работы!