КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Походження життя
|
|
|
|
Розглянемо основні події, які могли сприяти виникненню життя на нашій планеті, в аспекті гіпотези абіогенезу Опаріна-Холдейна.
Сучасний погляд на утворення Сонячної системи та місце Землі серед планет. Сучасна астрономія стверджує, що Земля та інші планети Сонячної системи утворилися з газово-пилової речовини близько 5 млрд років тому. Така газово-пилова матерія зустрічається в міжзоряному просторі й у наш час. Щоб на планеті могло виникнути життя, вона повинна мати певні розміри та одержувати енергію від якої-небудь зірки.
Розміри планети. Маса планети не повинна бути занадто великою, тому що енергія атомного розпаду природних радіоактивних речовин може спричинити перегрівання планети або, що ще більш важливо, радіоактивне забруднення середовища, що несумісно із життям. У той же час у занадто маленьких планет є інший недолік. Вони не можуть утримувати біля себе атмосферу, тому що сила тяжіння в них невелика (наприклад, Місяць).
Джерело енергії. Для виникнення життя і його подальшого існування необхідна енергія. Постійно й рівномірно одержувати необхідну кількість енергії дозволяє рух планети навколо якої-небудь зірки по коловій чи близькій до колової орбіті. Крім того, необхідна постійна інтенсивність випромінювання світила. Це дуже важливо, тому що інакше потік променистої енергії, який надходить на планету, не буде рівномірним. Нерівномірність потоку енергії призведе до різких коливань температури, а це перешкоджатиме виникненню й розвитку життя, тому що існування живих організмів можливе в певному температурному діапазоні (адже все живе на 70-80 % складається з води). Усім цим вимогам відповідала планета Земля.
Отже, близько 5 млрд років тому на Землі виникли космічні, планетарні й хімічні умови для більш високого рівня розвитку матерії — її еволюції в напрямку виникнення життя.
Основні етапи хімічної еволюції, яка передувала абіогенезу. Ці етапи включають утворення хімічних елементів і найпростіших неорганічних сполук.
Утворення хімічних елементів. Утворення хімічних елементів у процесі розвитку зоряних систем, у тому числі і таких, як наша Сонячна система, — закономірне явище в еволюції матерії.
Завдяки спектральному аналізу доведено, що водень є найбільш поширеним елементом у Всесвіті. Унаслідок реакцій ядерного синтезу з нього виникає гелій, з якого, у свою чергу, утворюється вуглець. У результаті приєднання до ядра вуглецю інших ядер гелію виникають ізотопи кисню, неону, магнію й інших елементів. Таким чином, виникнення атомів хімічних елементів, з яких складається основна маса зірок, планет і їх атмосфер, є лише початковим етапом неорганічної еволюції. Однак для подальшого розвитку в напрямку до виникнення життя необхідні були певні космічні й планетарні умови.
Утворення найпростіших неорганічних речовин. Водень, вуглець, кисень, азот і фосфор (біогенні елементи) досить поширені в космосі і можуть вступати в реакції, утворюючи при цьому найпростіші неорганічні сполуки, — це наступний етап неорганічної еволюції. Сприятливим чинником є надходження енергії від зірок у вигляді електромагнітного випромінювання й тепла.
Початковий етап існування Землі відзначався інтенсивним термоядерним процесом, високою температурою (понад 1000 °С) та активною хімічною діяльністю. У міру охолодження планети важкі елементи переміщалися до її центра, а більш легкі сполуки (H2, CO2, CH4та ін.) залишалися на поверхні. Метали й інші елементи, здатні окислюватися, з'єднувалися з киснем, і в атмосфері планети вільного кисню не було. Уся атмосфера ранньої Землі складалася з вільного водню і його сполук, тому мала відновний характер. Саме ця атмосфера — не окисна, а відновна — і сприяла виникненню життя. Вона складалася з водяної пари (Н2O), метану (СН,), діоксиду вуглецю (С02), оксиду вуглецю (СО), аміаку (NH3), азоту (N2), сірководню (H2S) та ін.
Останні спостереження радіоастрономії міжзоряного простору показали, що ті самі молекули, які виникають одночасно з народженням зірок, присутні в первинній атмосфері будь-якої планети. Багатьом радіоастрономам удалося ідентифікувати в міжзоряному просторі аміак і водяну пару.
Відповідні експерименти, проведені в той же час, підтвердили, що за наявності метану, води й аміаку напевно можуть утворитися такі сполуки, як ціаністий водень (HCN) і формальдегід (НСОН). Саме ці молекули і було виявлено в міжзоряному просторі. Сам факт існування таких сполук свідчить про те, що вони могли виникнути з первинних газових сумішей у результаті реакцій, пов'язаних з формуванням нових зірок. Це підготувало умови для третього етапу еволюції, який безпосередньо й започаткував процес абіогенезу.
Абіогенез. Перший етап — утворення простих органічних сполук. Для синтезу перших "цеглинок" життя необхідна була "сировина", тобто "будівельний матеріал" і енергія.
Сировиною могли бути N2 Н2, CO, СO2, Н20, NH3, СН4, H2S, HCN, НСОН та ін.
Джерелами енергії були: теплова енергія — тепло земної кори, яка охолоджувалася; а також енергія вулканічних вивержень і метеоритів, що потрапляли в атмосферу Землі; іонізуюча радіація — космічні промені й енергія радіоактивного розпаду важких елементів; сонячне світло — у першу чергу могутнє ультрафіолетове випромінювання, яке за відсутності озонового екрана безперешкодно проникало на Землю; енергія електричних розрядів від ударів блискавок; ударні хвилі (унаслідок падіння метеоритів).
Про цей етап, тобто про утворення мономерів — "цеглинок" біологічних полімерів — з газів, що містилися в первинній атмосфері, відомо найбільше, тому що ці реакції можна відтворити й повністю змодел ювати в лабораторних умовах. Досить успішні експерименти в цій галузі було проведено вже в 1913 році (Ж. Леб). Але найбільшу популярність здобули експерименти американських біохіміків С. Міллера і Юрі, які в 1953 р. переконливо довели можливість таких перетворень. Пропускаючи протягом декількох днів через суміш газів Н2, СН4 і NH3 і водяної пари електричний розряд при напрузі 60 тис Вольт (що за кількістю енергії можна порівняти з тривалістю періоду близько 50 млн. років на примітивній Землі) вони одержали у водяній фазі суміш, що складалася з різноманітних органічних речовин, у тому числі із 6 амінокислот (серед них — аспарагін; гліцин, глутамін), 11 органічних кислот — від мурашиної й оцтової до бурштинової, сечовину і метилсечовину.
Надалі виявилося, що абіогенним шляхом за умови відсутності кисню з отруйної суміші газів (N2 Н2, CO, СO2, NH3, СН4, H2S, HCN, НСОН) і водяної пари (Н,0) за участі розчинів солей важких металів, суміші глин та мінералів як каталізаторів, а як джерела енергії — електричних розрядів, ультрафіолетового випромінювання, іонізуючої радіації і дуже високої температури, можна синтезувати дуже багато простих органічних сполук, які входять до складу біологічних полімерів: білків, полісахаридів, ліпідів та нуклеїнових кислот.
Так, американський біохімік С. Фокс синтезував майже всі життєво важливі амінокислоти.
Етапи синтезу амінокислот, які утворюються під дією електричного розряду або ультрафіолетового випромінювання, якщо їх пропускати через суміш газів Н2, СН4 і NH,i водяну пару, досліджували у свій час Міллер і Юрі. Вивчаючи появу й зникнення окремих продуктів синтезу, що тривав протягом багатьох тижнів, вони переконалися, що концентрація аміаку неухильно знижувалася, а азот аміаку входив до складу насамперед щаністого водню (HCN) і ціаногену (C2N2). Це і були поряд з альдегідами перші синтезовані сполуки. Амінокислоти синтезувалися пізніше з ціаністого водню й альдегідів. Такий хід синтезу дозволяє припустити, що амінокислоти утворювалися з альдегідів шляхом, який добре відомий хімікам-органікам під назвою реакції Штрекера.
Крім амінокислот, при нагріванні газових сумішей в закритих посудинах до високих температур (600-900 °С) за участі руд різних металів як каталізаторів утворювалися жирні кислоти й деякі цукри. Серед синтезованих штучним шляхом цукрів є шести-вуглецеві (гексози) і п'ятивуглецеві (пентози). Рибозу й дезоксирибозу, зокрема, було отримано в результаті впливу на суміш газів ультрафіолету і гамма-випромінювання.
Ще Бутлеров у XIX столітті довів, що при нагріванні в міцному лужному розчині формальдегід (НСОН) перетворюється на суміш цукрів.
У XX столітті інші автори (Поннамперум і Бейбел) запропонували модель виникнення цукрів під дією гідротермальних джерел. Пропускання розведеного розчину формальдегіду над поверхнею нагрітої глини або навіть звичайне кип'ятіння формальдегіду спричинювали реакції конденсації та утворення цукрів.
У наш час хіміки-органіки досить легко одержують цукри із суміші метану (СН.), аміаку (NH3) і Н20 завдяки дії іонізуючої радіації. При цьому пентози, зокрема, утворюються в результаті конденсації проміжного продукту — формальдегіду (НСОН). Цей процес складається з ряду етапів, але сумарна реакція проста: із 5-ти молекул формальдегіду утворюється одна молекула рибози.
Як відомо, гексози (глюкоза, зокрема) входять як мономери до складу полісахаридів, а пентози (рибоза і дезоксирибоза) — до складу нуклеотидів, які є мономерами нуклеїнових кислот. Для утворення нуклеотида, крім пентози, потрібні ще фосфорна кислота й одна з азотистих основ, які належать до двох класів — пуринів і піримідинів.
Неодноразово робилися спроби — і досить успішні — продемонструвати можливість абіогенного синтезу азотистих основ, які входять до складу нуклеїнових кислот, шляхом ряду реакцій конденсації із ціаністого водню (HCN), який, у свою чергу, вдається одержати при нагріванні до високих температур сумішей з метану (СН4), аміаку (NH3) і води.
Ціаністий водень (HCN), який є смертельною отрутою для сучасної біосфери, на первинній Землі, імовірно, був дуже поширеною сполукою і віді гра вав важливу роль в абіогенних процесах. Результати спектроскопічних досліджень свідчать про те, що він входить і до складу комет. Згідно з останніми радіоастрономічними даними, HCN поширений також у міжзоряному просторі.
Завдяки різноманітним реакціям, джерелом яких був HCN, його похідні і деякі інші сполуки, в умовах, що наближалися до умов первісної Землі, було отримано аденін, гуанін, урацил і цитозин. При цьому найлегше синтезується аденін.
Одним з перших досягнень у синтезі пуринів у можливих первинних умовах Землі була робота Хуана Оро з університету Х'юстона, який шляхом термічного впливу на ціаністий амоній одержав аденін.
Якщо уважно розглянути молекулу аденін а, про склад якої дає уявлення емпірична формула C5H5N5, можна встановити, що він являє собою пентамер ціаністого водню, тобто з 5-й молекул HCN утворюється одна молекула C5H5N5. Ймовірно, що спочатку з 4-х молекул ціаністого водню (HCN) виникає його тетрамер C4H4N4 (діаміномалеонітрил). Ця сполука є важливим проміжним продуктом у багатьох реакціях, що приводять до синтезу азотистих основ. Виявилося, що під дією світла може відбуватися перебудова молекули C4H4N4 із приєднанням ще однієї молекули HCN, у результаті чого утворюється аденін.
Трохи іншим шляхом можна одержати іншу азотисту основу, яка входить до складу нуклеотидів, — гуанін. Вихідним продуктом для його синтезу є той же діаміно-малеонітрил (C4H4N4). Але в даному випадку вдаються до реакції гідролізу за участю ціаногена (C2N2).
Складніше із синтезом піримідинів (тиміну, урацилу і цитозину), однак певного успіху досягнуто й тут. Так, Оро вдалося синтезувати урацил шляхом взаємодії аміачного розчину сечовини зі сполуками, які виникають із простих газів під впливом електричних розрядів. Л. Оргель синтезував цитозин, нагріваючи разом сечовину і ціаноацетилен (C3HN), який утворюється, у свою чергу, у результаті впливу електричного розряду на суміш метану й азоту.
Азотисті основи (аденін, гуанін, тимін, урацил і цитозин), з'єднуючись з пентозами (рибозою або дезоксирибозою), утворюють нуклеозиди. Останні, у свою чергу, з'єднуючись з фосфорною кислотою, утворюють нуклеотиди. У результаті приєднання аденіну до рибози утворюється аденозин. Із нього після приєднання до молекули "хвоста", який складається із трьох фосфатних груп, утворюється аденозинтрифосфат (АТФ).
Слід зауважити, що для поєднання із трифосфатом у процесі абіогенезу було вибрано саме аденозин, а не який-небудь інший нуклеозид. Немає ніяких явних вказівок на те, що АТФ пристосований для збереження енергії краще, ніж ГТФ (гуанозинтри-фосфат), ЦТФ (цитидинтрифосфат), ТТФ (тимідинтрифосфат) чи УТФ (уридин-трифосфат). Можливо, що аденін використовувався для синтезу енергозберігаючої сполуки саме тому, що він був компонентом первинного "бульйону" у більш високій концентрації, оскільки синтез його відносно простий.
Підбиваючи підсумок, можна сказати, що подібні результати, які підтверджують теорію абіогенного утворення найпростіших органічних сполук, були отримані в усіх вищенаведених та інших експериментах. При цьому виявилося, що із суміші газів (N, Н2, CO, CO., NH3, СН4, H2S) і водяної пари спершу утворяться високоактивні проміжні сполуки, такі, як ціаністий водень (HCN), формальдегід (НСОН), ціаноген (CjNj) та ін., а вже потім із цих сполук утворяться в результаті хімічної еволюції мономери біологічних полімерів; саме вони і становили вміст "первинного бульйону" на нашій планеті. Звідси випливає, що виникнення "первинного бульйону" — не випадковість, а закономірність. На планеті з оптимальною температурою, достатньою кількістю води та атмосферою, що складається із найбільш розповсюджених у Всесвіті газів, "первинний бульйон" повинен був виникати в 100 випадках зі 100.
На цьому закінчується 1-й етап хімічної еволюції. Наступна фаза — утворення полімерів, оскільки амінокислоти, жирні кислоти, цукри та азотисті основи — це ще не життя.
Другий етап — утворення біополімерів. Наступним кроком еволюції було утворення з малих органічних мономерів крупніших — полімерів, схожих на білки та нуклеїнові кислоти.
Білкові молекули й молекули нуклеїнових кислот — найскладніші з біологічних полімерів. їх синтез у клітині являє собою багатоступінчастий ферментативний процес, який відбувається зі споживанням енергії. Джерелом енергії є АТФ (або ГТФ у деяких реакціях синтезу білка). Навряд чи можна вважати ймовірною раптову появу в процесі хімічної еволюції одиничної молекули нуклеїнової кислоти або таку ж появу молекули білка. Тільки сполуки, які виникали у великих кількостях, могли стати основою для подальшого розвитку. Крім того, ці сполуки звичайно ж не були настільки складними, як сучасні білки й нуклеїнові кислоти. Поява великих органічних молекул — важлива подія в хімічній еволюції, що передувала появі життя на Землі, — була, очевидно, також багатоступінчастим процесом.
Аналіз цього етапу в експериментальному й виглядає набагато бідніше, хоч і тут є деякі моделі. Цей цикл експериментів пов'язаний з конденсацією або полімеризацією мономерів з метою одержання більш крупних молекул. Як відомо, ці реакції відбуваються з виділенням води, тобто супроводжуються дегідратацією. Так, наприклад, при поєднанні (конденсації) амінокислот у поліпептидний ланцюжок (первинна структура білкової молекули) за місцем утворення пептидного зв'язку (зв'язок між карбоксильною групою й аміногрупою) відбувається виділення молекули води. Коли утворюється нуклеїнова кислота, то відбувається аналогічний процес: фосфат з'єднується із цукрами, а цукри — з основою, при цьому виділяється молекула води. Оскільки для синтезу полімеру необхідно швидше видалити воду, то, імовірно, найлегше це зробити шляхом нагрівання суміші амінокислот. Такий же процес можливий і при утворенні нуклеїнової кислоти: суміш основи й цукрів нагрівається, що спричинює їх дегідратацію і приводить до утворення нуклеозиду. Подібним же чином, нагріваючи нуклеозид і фосфат, можна одержати нуклеотид.
Для первинної Землі така послідовність подій цілком імовірна. Першим, хто звернув на це увагу, був англійський фізик Дж. Бернал. На його думку, морські лагуни, що періодично пересихають, могли бути ідеальним місцем для утворення великих молекул. Вплив сонячного випромінювання на органічний матеріал, адсорбований на мулистому дні лагун, призводив до його дегідратації. Полімери, які виникали при цьому, змивалися потім водою в океан. Подібні реакції могли відбуватися вздовж усієї прибережної смуги.
Одна з найбільш вдалих моделей цього етапу належить С. Фоксу, який у результаті нагрівання суміші сухих амінокислот одержав поліпептиди різної довжини. Вони дістали назву "протеїноїдів", тобто попередників білків. Отримані поліпептиди можна було потім розчиняти у воді; при цьому вони залишалися стабільними. В одному з експериментів Фокс замість колби використав видовбаний шматок вулканічної лави. Вихід пептидів при цьому досягав 40 % від початкової кількості амінокислот. На переконання Фокса, центрами виникнення життя на цьому етапі були гарячі, сухі вулканічні райони, яких на той час на Землі було багато. Гарячі схили вулканів покривалися кіркою органічних речовин; дощ змивав пептиди, які утворювалися, в океан. Навряд чи первинний океан нагадував сучасний, якщо порівнювати глибини. Води на первинній Землі було, швидше за все, мало, такий океан могла б перебрести і курка. Саме незначна кількість води забезпечила високу концентрацію низькомолекулярних речовин у "первинному бульйоні" і зробила ймовірною їх взаємодію.
Якщо для утворення простих органічних речовин необхідні були досить могутні джерела енергії, то для утворення полімерів достатньо було простого нагрівання. Однак для реакції полімеризації необхідні каталізатори. У клітині цю роль виконують ферменти. Було висловлено припущення, що каталізаторами в процесі синтезу полімерів на первинній Землі могли бути поверхні мінеральних глин. Експериментально було доведено, що розчин амінокислоти аланіну у водному середовищі за присутності особливого виду глинозему й АТФ може давати полімерні ланцюжки поліаланіну.
Таким чином на давній Землі могли утворитися поліпептиди. Деякі з них були, можливо, каталітично активними. Однак це могло і не мати безпосереднього стосунку до виникнення життя, оскільки поліпептиди не мають здатності до самовідтворення, що є найбільш важливою ознакою живого. Ця властивість є, як відомо, у нуклеїнових кислот, які здатні до реплікації.
Були спроби синтезувати нуклеїнові кислоти неферментативним способом. Вихідною речовиною для цього були високоенергетичні фосфати. Завдяки цим сполукам вдалося одержати полінуклеотиди навіть при помірних температурах. Деякі реакції проводилися при температурі 150-160 °С, але полімеризація відбувалася навіть при температурі 65 °С.
Цікавими виявилися експерименти, коли в середовищі, що містить солі фосфорної кислоти (фосфати) і рибонуклеотиди у високих концентраціях, спонтанно синтезуються короткі полірибонуклеотиди. Також спонтанно, шляхом спарювання комплементарних основ, можуть утворитися РНК-копії. Обидві ці реакції протікають без участі ферментів чи інших білків. Звідси випливало, що РНК може виявляти власну каталітичну активність, що й підтвердив американський біохімік Т. Чек, який відкрив у 1982 р. каталітичні властивості РНК.
Поки що важко встановити, як могла виникнути ДНК. Ця органічна речовина пристосована краще, ніж РНК, до довгострокового зберігання інформації. В усякому разі відомо, що відсутність кисню в 2-положенні дезоксирибози робить молекулу ДНК більш стійкою, на відміну від РНК, до гідролітичного розщеплення в слаболужних водних розчинах, а саме такі розчини були в первинних водоймах і збереглися в сучасних клітинах. Крім того, наявність двох комплементарних ланцюгів ДНК забезпечує процес реплікації і полегшує виправлення помилок, які виникають у кожному із двох ланцюжків ДНК. Цілком можливо, що завдяки активності давнього білка, який є близьким до сучасного фермента — зворотної транскриптази (тобто здатний синтезувати ДНК, використовуючи як матрицю РНК), і утворилися перші молекули ДНК на Землі.
Незрозумілим залишається також питання про те, як найдавніша РНК почала кодувати амінокислотну послідовність. Але коли це відбулося, то утворилися системи зі зворотним зв'язком, у яких нуклеїнові кислоти містили інформацію про структуру білків, що сприяло збільшенню кількості кодуючих їх нуклеїнових кислот. Крім того, білки могли захищати нуклеїнові кислоти від руйнівної дії ультрафіолету.
Обговорюється в літературі і питання про те, що ж з'явилося на Землі раніше: поліпептидна послідовність чи полінуклеотидна, тобто що було первинним — білок чи нуклеїнова кислота?
Генетичну гіпотезу, або гіпотезу "голого гена", запропонували ще в 1929 р. Дж. Холдейн і Г. Мюллер. Відповідно до цієї гіпотези, яку підтримали згодом й інші автори, спочатку виникли нуклеїнові кислоти як матрична основа для синтезу більш пізніх за часом утворення білків. Доказом на користь цього факту можна вважати відкриття каталітичної властивості РНК.
З наведених даних зрозуміло, що в цій гіпотезі, коли йшлося про процеси абіогенезу, данина віддавалася пріоритету такої властивості живого, як здатності до самовідтворення. О. І. Опарін, який першим запропонував гіпотезу абіогенезу (1924 p.), спочатку припускав, що первинним є білок, оскільки без ферментів (тобто білків) неможливий обмін речовин, а без останнього неможливі ніякі прояви життя. Цілком очевидно, що в цій гіпотезі віддається данина пріоритету в процесах абіогенезу такій важливій властивості живого, як обмін речовин.
Однак після того, як з'явилися відомості про будову й роль нуклеїнових кислот у проявах життя, О. І. Опарін дещо змінив свої погляди і в останній своїй монографії "Матерія - життя - інтелект" (1977 р.) висловив чудову думку про те, що поліпептиди й полінуклеотиди ще на зорі життя на Землі еволюціонували разом шляхом взаємодії, спочатку далеко ще не зовсім досконалої, тобто відбувалася спільна еволюція, або коеволюція.
Таким чином, об'єднання здатності до самовідтворення полінуклеотидів з каталітичною активністю поліпептидів слід вважати однією з найбільш важливих сходинок хімічної еволюції. Для виникнення життя необхідною була участь як полінуклеотидів, так і поліпептидів. Найбільші шанси на збереження в ході передбіологічного добору мали ті системи, у яких здатність до обміну речовин поєднувалася зі здатністю до самовідтворення.
У подальшому ускладненні обміну речовин у таких системах істотну роль повинні були відігравати каталізатори й просторово-часове роз'єднання початкових і кінцевих продуктів реакції. Останнє не могло виникнути без фазово-контрастного поділу, тобто без виникнення мембрани й утворення, яке передувало клітині, — у цьому й полягає основна сутність наступного етапу еволюції, про який ми також можемо розмірковувати лише гіпотетично.
Третій етап — виникнення пробіонтів і біологічних мембран. Питання про те, як відбувався перехід від біополімерів до перших живих істот, є найскладнішим моментом у проблемі виникнення життя. Найбільш відомий підхід до вирішення цього питання міститься в експериментах О. І. Опаріна, який запропонував коацерватну гіпотезу (1924 p.).
О. І. Опарін припустив, що перехід від хімічної еволюції до біологічної пов'язаний з виникненням найпростіших фазово-відособлених органічних систем — пробіонтів, здатних використовувати речовину й енергію з навколишнього середовища і завдяки цьому здійснювати найважливішу життєву функцію — функцію росту.
Найбільш вдалим об'єктом для моделювання виявилися коацерватні краплі (коацервати). О. І. Опарін спостерігав, як у колоїдних розчинах поліпептидів, полісахаридів, РНК та інших високомолекулярних сполук за певних умов утворюються згущення, що мають об'єм від 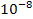 до
до 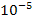 см3. Ці згущення і є коацерватами. Навколо коацерватних крапель є межа розділу, яку можна побачити в мікроскоп. Коацервати здатні адсорбувати речовини із зовнішнього середовища, у них усередині вже може здійснюватися синтез нових сполук. Оскільки концентрація речовин у коацерватній краплі більша, ніж у навколишньому середовищі, то це означає, що і хімічні реакції відбуваються з більшою швидкістю. Досягаючи певного розміру, під дією механічних сил коацерватні краплі можуть поділятися на менші частини. Але коацервати — неживі істоти. Це лише найпростіші моделі пробіонтів, що виявляють зовнішню подібність з такими властивостями живого, як ріст та обмін речовин з навколишнім середовищем.
см3. Ці згущення і є коацерватами. Навколо коацерватних крапель є межа розділу, яку можна побачити в мікроскоп. Коацервати здатні адсорбувати речовини із зовнішнього середовища, у них усередині вже може здійснюватися синтез нових сполук. Оскільки концентрація речовин у коацерватній краплі більша, ніж у навколишньому середовищі, то це означає, що і хімічні реакції відбуваються з більшою швидкістю. Досягаючи певного розміру, під дією механічних сил коацерватні краплі можуть поділятися на менші частини. Але коацервати — неживі істоти. Це лише найпростіші моделі пробіонтів, що виявляють зовнішню подібність з такими властивостями живого, як ріст та обмін речовин з навколишнім середовищем.
Особливе значення в еволюції пробіонтів мало формування каталітичних систем. Першими каталізаторами були неорганічні сполуки, такі як солі заліза, міді й інших важких металів. Однак їх дія була досить слабкою. Поступово на основі передбіологічного добору виникли біологічні каталізатори, що мало велике значення для удосконалення обміну речовин.
Справжній початок біологічної еволюції ознаменувало виникнення пробіонтів з кодовими взаємозв'язками між білками і нуклеїновими кислотами. Взаємодія білків і нуклеїнових кислот зумовила виникнення таких властивостей живого, як здатність до самовідтворення, до збереження спадкової інформації і передавання її наступним поколінням. Пробіонти, у яких обмін речовин поєднувався зі здатністю до самовідтворення, мали найкращу перспективу зберегтися в передбіологічному доборі.
Інший механізм утворення клітин запропонував С. Фокс з університету в Ма-йамі. Як уже зазначалося, Фокс шляхом нагрівання безводної суміші амінокислот одержував поліпептиди. Учений назвав їх протеїноїдами. У результаті розчинення протеїноїдів у гарячій воді він одержав рідину — непрозору суспензію. Під мікроскопом вдалося побачити, що ця суспензія складається з крихітних кульок, які нагадують сучасних бактерій-коків. Фокс назвав їх мікросферами, тобто маленькими кульками. Деякі з них утворювали ланцюжки або були схожі на дріжджові клітини в період поділу. Електронний мікроскоп показав, що кожна мікросфера оточена подвійною клітинною оболонкою, дуже схожою на мембрани сучасних клітин. Надалі Фокс проводив досліди з комплексами протеїноїдів і полінуклеотидів. Ці комплекси також могли утворювати мікросфери, причому такі, що могли ділитися й брунькуватися. їх подібність до сучасних мікроорганізмів вражає.
Яка з моделей ближча до п рото клітини — коацервати Опарі на чи мікросфери Фокса — важко сказати. У всякому разі, у передбіологічному доборі лише ті утворення виявилися конкурентоздатними, які оточили себе зовнішньою мембраною.
Біологічні мембрани, як відомо, складаються з білків і ліпідів. Вони відокремлюють речовину від середовища й надають міцності молекулярному агрегату. Є дві найбільш розповсюджені точки зору з приводу можливого виникнення біологічних мембран. Відповідно до однієї з них, мембрани могли виникнути в ході формування коацерватів, які утворювалися у воді при зіткненні двох слабо взаємодіючих полімерів. Відповідно до іншої точки зору, утворення мембран відбувалося при адсорбції полімерів на поверхні глин. Поки що немає достатніх підстав, щоб віддати перевагу якійсь одній із цих двох точок зору.
Поява біологічної мембрани, яка відокремлює вміст пробіонта від навколишнього водного середовища і має здатність до вибіркової проникності, визначила напрямок подальшої хімічної еволюції середовища по шляху розвитку все більш досконалих, здатних до саморегулювання систем — аж до виникнення перших примітивних клітин. Подальший їх розвиток уже повністю набуває рис біологічної еволюції, яка охоплює проміжок часу не менш як 3,5 млрд. років.
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 402; Нарушение авторских прав?; Мы поможем в написании вашей работы!