КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вуглецевий
|
|
|
|
цикл — кругообіг вуглецю між атмосферою, океаном, біосферою та надрами Землі.
Колообіг вуглецю у природі включає біологічний цикл, виділення СО2 в атмосферу при згорянні палива, із вулканічних газів, гарячих мінеральних джерел, із поверхневих шарів океанічних вод та ін.

Біологічний колообіг
Біологічний колообіг вуглецю пов’язаний з життєдіяльністю організмів. Біологічний цикл полягає в тому, що вуглець у вигляді СО2 поглинається із тропосфери рослинами. Потім із біосфери знову повертається в геосферу: з рослинами вуглець потрапляє до організму тварин та людини, а потім при гнитті тваринних та рослинних матеріалів – до ґрунту, і у вигляді СО2 – до атмосфери.
Вуглець – основний біогенний елемент. Він відіграє важливу роль в утворенні живої речовини біосфери. Вуглекислий газ із атмосфери в процесі фотосинтезу, який здійснюють зелені рослини, асимілюється і перетворюється на численні різноманітні органічні сполуки рослин. Рослинні організми, особливо нижчі мікроорганізми, морський фітопланктон, завдяки виключній швидкості розмноження, виробляють на рік близько 1,5• 107 т вуглецю у вигляді органічної маси.
Рослини часто поїдаються тваринами. При цьому утворюються більш або менш складні трофічні зв’язки. В остаточному підсумку органічна речовина в результаті дихання організмів, розкладу їхніх трупів, процесів бродіння, гниття та горіння перетворюється на вуглекислий газ або відкладається у вигляді сапропелю, гумусу, торфу, які, в свою чергу, дають початок багатьом іншим каустобіолітам – кам’яному вугіллю, нафті, горючим газам. Біологічний цикл вуглецю закінчується або окисненням і поверненням у атмосферу, або відкладенням у вигляді вугілля або нафти.
В процесах розпаду органічних речовин величезну роль відіграють бактерії та гриби. В активному колообігу вуглецю бере участь дуже невелика частка всієї його маси. Величезну кількість вугільної кислоти законсервовано у вигляді вапняків та інших порід. Між вуглекислим газом атмосфери і води океану, в свою чергу, існує рухома рівновага.
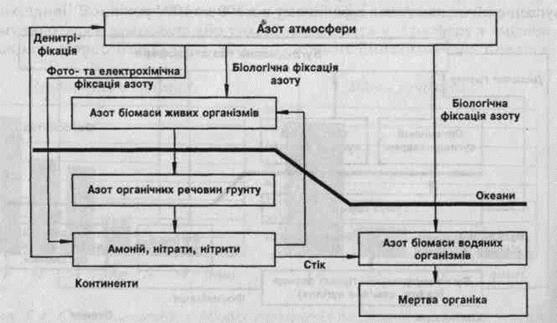
Кругообіг Нітрогену в пироді
Хімічні елементи, як і вся природа, перебувають у постійному русі. Процеси відбуваються в усіх трьох оболонках Землі — літосфері, гідросфері, атмосфері. Значну роль у процесах, що відбуваються у природі, обирає біосфера — зона існування живих організмів. Так, вам відомо, що Нітроген входить до складу білків і, отже, зумовлює існування рослин, тварин, взагалі життя на 3eмлі.
У природі Нітроген зустрічається як у вільному стані, так і у зв'язаному. У вільному стані Нітроген у вигляді азоту входить до складу повітря (об'ємна частка N2 становить 78 %, масова — 75,6 %). Оскільки азот з повітря витрачається мало, його запаси в атмосфері залишаються сталими. У вигляді неорганічних сполук Нітроген у невеликих кількостях є в ґрунті. Проте у вигляді складних органічних сполук — білків — він увіходить до складу всіх живих організмів, беручи участь у їх життєдіяльності.
З Нітрогену складається азот N2, його багато в повітрі. Проте безпосередньо з повітря Нітроген у вигляді азоту засвоюють лише деякі бактерії, а всі інші організми здатні засвоювати Нітроген тільки у складі сполук.
Рослини засвоюють Нітроген неорганічних сполук, які у ґрунті, у вигляді йонів NH4 і NO3. У рослинах здійснюється синтез білків. Рослини частково поїдаються травоїдними тваринами, і білкові речовини потрапляють до організму тварин. Під час гниття залишків рослин і тварин під впливом спеціальних бактерій відбуваються складні біохімічні процеси, внаслідок яких органічні сполуки, що містять Нітроген, перетворюються на неорганічні сполуки Нітрогену, які повертаються в ґрунт. Ці сполуки знову засвоюються рослинами, і цикл перетворень замикається.
2. Екологічні групи рослин по відношенню до води.
1. Гідатофіти – це водяні рослини, цілком або майже цілком занурені у воду. Вийняті із води, ці рослини швидко висихають і гинуть, бо в них редуковані продихи і нема кутикули. До них належать водяний жовтець, елодея, валіснерія, рдесник, водопериця.
2. Гідрофіти – це наземно-водяні рослини, частково занурені у воду.Ростуть на берегах водойм, на мілководді та на болотах. До них відносять очерет звичайний, частуху подорожникову, бобівник трилистий, калюжницю болотяну. У них краще, ніж у гідатофітів розвинуті механічні та провідні тканини.
3. Гігрофіти – це наземні рослини, які живуть в умовах підвищеної вологості повітря або на досить вологих грунтах. Серед них розрізняють тіньові і світлові гігрофіти.
Тіньові – це рослини нижніх ярусів сирих лісів у різних кліматичних зонах. До них належать будяк городній, розрив-трава, цірцея альпійська, тропічні трави.
Світлові гігрофіти – це види відкритих місцевостей помірної смуги, які ростуть на постійно вологих грунтах і у вологому повітрі – рис, підмаренник, росичка, папірус.
4. Мезофіти – це рослини, що можуть переносити нетривалу і не дуже сильну посуху. Вони ростуть при середній вологості, помірно теплому режимі і досить добрій забезпеченості мінеральним живленням. До мезофітів відносять вічнозелені дерева верхніх ярусів тропічних лісів, листопадні дерева саван, деревні породи вологих вічнозелених субтропічних лісів,чагарники підліска, трав’янисті рослини дібров, рослини заплавних і не надто сухих лук, багато бур’янів і більшість культурних рослин. Отже, група мезофітів дуже численна і неоднорідна.
5. Ксерофіти – це рослини, що ростуть у місцях з недостатнім зволоженням і мають пристосування, які дають змогу добувати воду в разі її нестачі або обмежувати її випаровуванні і, навіть, вміння запасати її під час посухи. Ксерофіти краще ніж усі інші рослини здатні регулювати водний обмін, тому під час тривалої посухи перебувають в активному стані. Ксерофіти поділяються на дві основні групи: сукуленти і склерофіти.
Сукуленти – соковиті рослини з дуже розвинутою водозапасаючою паренхімою в різних органах. Тому сукуленти є:
-стеблові–кактуси,кактусовиднімолочаї,стапелії;
-листкові–алое,агави,бріофілуми,молодило,очитки;
-кореневі–аспарагус,квасениця.
Склерофіти – рослини, сухі на вигляд, часто з вузькими та дрібними листками, іноді скрученими в трубочки.
Таким чином, неоднакові шляхи регуляції водного обміну дають змогу рослинам заселяти різні за екологічними умовами ділянки суші. Різноманітність пристосувань лежить в основі поширення рослин по поверхні планети, де дефіцит вологи є однією з головних проблем екологічних адаптацій.
3. Обмін речовин
Обмін речовин або метаболізм — сукупність хімічних реакцій, що відбуваються в живих організмах. Метаболізм поділяється на дві гілки: катаболізм (дисиміляція або енергетичний обмін), що включає реакції розщеплення складних органічних речовин до простіших, яке супроводжується їх окисненням і виділенням корисної енергії, та анаболізм(асиміляція або пластичний обмін) — реакції синтезу необхідних клітині речовин, у яких енергія, отримана у катаболічних реакціях, використовується.
Майже всі метаболічні реакції пришвидшуються ферментами —каталізаторами білкової природи. Ферменти не тільки роблять можливим швидке протікання у клітині великої кількості реакцій, що за інших умов потребували би дуже високих температур або/і тиску, а й дозволяють регулювати їх за потреби. Реакції каталізовані ферментами часто об'єднуються у послідовності, де продукт однієї стає субстратом для наступної, такі серії реакцій називаються метаболічними шляхами. Метаболічні шляхи в свою чергу поєднуються між собою, утворюючи складні розгалужені сітки.
Реакції катаболізму — це окиснення органічних речовин, тобто відщеплення від них електронів. Кінцевим акцептором цих електронів можуть виступати ендогенні органічні речовини, такий тип катаболізму називається бродінням переважно протікає за відсутності кисню і є основним шляхом отримання енергії для багатьох мікроорганізмів. Якщо у серії катаболічних реакцій кінцевим акцептором електронів є екзогенні речовини, то вона називається клітинним диханням. Дихання поділяється на аеробне, при якому акцептором виступає кисень, та анаеробне, акцепторами є інші речовини, переважно неорганічні.Усі типи клітинного дихання включають ланцюги транспорту електронів.
Анаболізм (асиміляція) — сукупність хімічних процесів в організмі, спрямованих на утворення та відновлення структурних компонентів органів і тканин; один з етапів проміжного обміну, що забезпечує ферментативний синтез складних сполук із простіших низькомолекулярних попередників за допомогою енергії АТФ та інших макроергічних сполук; тісно взаємопов’язаний із протилежним процесом — катаболізмом. Кінцеві продукти катаболізму, як правило, є вихідними продуктами анаболізму.
Ферменти або ензими — органічні каталізатори білкової або РНК природи, які утворюються в живих організмах, здатних прискорювати перебіг хімічних реакцій в організмі. Ферменти каталізують більшість хімічних реакцій, які відбуваються у живих організмах. Вони можуть мати від одного до кількох поліпептидних ланцюгів — субодиниць. Кожен із ферментів має один або більше активних центрів, які визначають специфічність хімічної реакції, що каталізується даним ферментом.
Всі біохімічні реакції відбуваються за участю ферментів за нормальним тиском, температурою, у слабокислому, нейтральному чи слаболужному середовищі.
Ферменти РНК-природи називаються рибозимами і вважаються первісною формою ферментів, які були замінені білковими ферментами в процесі еволюції.
4.Як отримують біологічно активні сполуки за допомогою ізольованих рослинних і тваринних тканин?
Головним напрямом біотехнології є виробництво з допомогою мікроорганізмів і культивованих эукаріотичних клітин біологічно активних сполук (ферменти, вітаміни, гормони), лікарських засобів і навіть цінних сполук.
Клітинна інженерія — це метод конструювання клітин нових типів з урахуванням їх культивування, гібридизації та реконструкції. Вона базована на використанні методів культури клітин та тканин.
Рослинні клітини у культурі — це джерело найцінніших природних речовин, оскільки вони зберігають здатність синтезувати притаманні їм речовини: алкалоїди, ефірні олії, смоли, біологічні активні сполуки. Так, перекладені на культуру клітини женьшеню продовжують синтезувати, як у складі цілісної рослини, і в результаті дають цінну лікарську сировину. Причому, у культурі з клітинами та його геномами можна робити будь-які маніпуляції. Використовуючи індукований мутагенез, можна підняти продуктивність штамів культивованих клітин та проводити їхню гібридизацію, що набагато легше й простіше, ніж на рівні цілісного організму.
Шляхом гібридизації лімфоцитів (клітин, синтезують антитіла, але неохоче, і недовго які у культурі) з пухлинними клітинами, з потенційним безсмертям та спроможними до необмеженому зростанню в штучної середовищі, вирішена одну з найважливіших завдань біотехнології на етапі — отримані клітини гібридоми, здатні до нескінченному синтезу высокоспецифических антитіл певного типу.
Отже, клітинна інженерія дозволяє конструювати клітини нових типів з допомогою мутаційного процесу, гібридизації і більше, комбінувати окремі фрагменти різних клітин (ядра, мітохондрії, пластиды, цитоплазму, хромосоми тощо. буд.), клітини різних видів, що стосуються як до найрізноманітніших пологам, сімействам, а й царствам. Це полегшує рішення багатьох теоретичних труднощів і має практичного значення.
8.Фотоперіодизм
Фотоперіоди́зм — фізіологічна реакція організмів на добовий ритм освітлення (співвідношення довжини дня та ночі). Зустрічається у рослин і тварин. Виявляється в коливаннях інтенсивності фізіологічних процесів. Найбільшою мірою фотоперіодизм властивий зеленим рослинам, життєдіяльність яких безпосередньо залежить від світлової енергії Сонця.
Фотоперіодизм відкрито в 1920 р. американськими вченими В. Гарнером і Г. Аллардом на прикладі тютюну.
У рослин
Фотоперіодизм у рослин проявляється в зміні процесів росту та розвитку. Один з основних проявів цієї реакції – фотоперіодична індукція цвітіння.
За типами ФПР розрізняють такі основні групи рослин (Т. К. Горишина, 1980):
Рослини короткого дня, яким для переходу до цвітіння потрібно світла 12 і менше годин на добу (коноплі, тютюн);
Рослини довгого дня, для фази цвітіння їм потрібна довжина дня понад 12 годин на добу (картопля, пшениця, шпинат);
Рослини проміжного типу, цвітіння в яких наступає при певному діапазоні фотоперіодизму, наприклад гваюла;
Рослини фотоперіодично нейтральні, цвітіння в яких відбувається при будь-якій довжині дня (помідор, кульбаба тощо).
Особливо велику роль фотоперіодизм відіграє в географічному поширенні рослин і в регуляції їх сезонного ритму.
Фотоперіодизм вивчали американські фізіологи рослин У. Гарнер, Г. Аллард у 1920-1925 рр. В залежності від реакції рослин на довжину дня, що прискорює зацвітання, вони поділяються на:
довгоденні (ДДР - рослини довгого дня – злаки, хрестоцвіті, кріп; поширені в основному в помірних та приполярних широтах);
короткоденні (КДР - рис, соя, конопля; субтропіки);
довгокороткоденні (ДКДР) і
короткодовгоденні (КДДР) – потребують чергування різних фотоперіодів;
нейтральні (НДР - гречка, горох і ін.).
Поділ рослин на дані групи не пов’язаний з конкретною оптимальною довжиною дня, а лише дає уявлення про те, чи прискорюється цвітіння при збільшенні чи зменшенні тривалості освітлення в кожному фотоперіоді.
Є рослини з якісними й кількісними типами фотоперіодичних реакцій. Інтенсивність освітлення, температура й інші фактори можуть змінювати характер фотоперіодичних проявів рослин.
Для КДР із якісною реакцією основним фактором служить тривалість темнового періоду. Його переривання (хоч на 1 хв.) перешкоджає переходу до цвітіння. Переривання світлового періоду темнотою не впливає на строки цвітіння. ДДР не потребують періоду темноти і зацвітають при безперервному освітленні.
Фотоперіодична дія сприймається головним чином листками, а не апексами пагонів. Максимальну чутливість до фотоперіоду мають листки, які тільки що закінчили ріст. Здійснюється це головним чином завдяки фітохрому. Вважають, що його темнове перетворення може служити способом вимірювання часу по типу піскового годинника. Спалах червоного світла встановлює фазу ендогенного ритму.
Необхідність великої поверхні листків і достатньої інтенсивності світла для переходу до цвітіння у багатьох рослин пояснюється потребою ростучих меристем в асимілятах. У НДР перехід до цвітіння забезпечується віковими змінами. Тут спрацьовує ендогенна регуляція. Дані фактори (температура і фотоперіод) можуть діяти послідовно, як, наприклад, в озимих злаків.
Температурна й фотоперіодична регуляція служать пристосуванням рослин до умов існування, тому що обумовлюють сприятливі строки для переходу до цвітіння. В ході фотоперіодичної індукції в листках утворюється стимулятор цвітіння, який транспортується у вегетативні бруньки пагонів, де включає другу фазу ініціації – евокацію.
У тварин
У тварин і людини фотоперіодізм виражається головним чином в коливаннях інтенсивності обміну речовин і енергії. У тварин з фотоперіодізмом зв'язані також терміни настання і припинення шлюбного періоду, плодючість, осінні і весняні ліньки, перехід до зимівлі, міграції і ін. За відношенням до світла тварини поділяються на групи в залежності від того, який спосіб життя вони ведуть: денні, нічні сутінкові.
13.Симбіоз.мутуалізм,паразитизм,антагонізм,коменсалізм.
(Приклади між рослиною і твариною,рослиною і м.о,бактеріями і людиною.)
Симбіоз - будь-які взаємозв’язки між організмами, основні види взаємозв’язків:
· антагонізм
· мутуалізм
· паразитизм
· коменсалізм
Антагоністичні відносини – це конкуретні відносини між м/о при якому відбуваються пригнічення розвитку одних організмів іншими. Приклад – нормальна мікрофлора організму і патогенні небезпечні організми. В людському організмі є багато корисних мікробів, зовні потрапляють патогенні мікроби і вони конкурують за середовище існування, за джерело живлення.
Мутуалізм – взаємосприятливі відносини.(Нітрифікуючі бактерії одні з них засвоюють аміак і утв токсичний нітрит, ін. група нітрифікуючих бактерій забирає їх і перетворює до нітрату.)
Паразитизм – тип взаємовідносин коли один з учасників чинить шкідливий вплив на м/о, захоплює клітини, тканини ін.. організму і використовує їх як середовище існування.
Коменсалізм – коли має вигоду тільки один учасник, а ін.. ні користі, ні шкоди.
11.ТИПИ ТРАНСГЕННИХ РОСЛИН ТА ЇХ ПОШИРЕННЯ.
Трансгенні рослини - це рослини, ДНК яких модифіковане шляхом застосування генетично інженерних методів. Основною ціллю створення ГМ рослин є представлення нових сортів із специфічними ознаками, які не притаманні для рослин цього виду. Прикладом таких ознак можуть бути стійкість до різного роду гербіцидів, шкідників, стійкість до несприятливих умов зовнішнього середовища (солестікість, засухостійкість тощо) чи набуття нових якостей харчового значення.
Є кілька досить поширених методів на впровадження чужорідної ДНК в геном рослини.
Метод 1:Існує бактерія Agrobacteriumtumefaciens. Під час інфікування відбувається вбудовування в геном рослинної клітини специфічного сегменту бактеріальної плазмідної ДНК - Т-ДНК. Т-ДНК - частина плазміди, яка індукує розвиток пухлини;
Метод 2:Клітини, попередньо оброблені спеціальними реагентами, які руйнують товсту клітинну оболонку, вміщують у розчин, у якому: ДНК і ті речовини, які б сприяли її проникненню у клітину.
Метод 3:Існує метод бомбардування рослинних клітин спеціальними, дуже маленькими вольфрамовыми кулями, що містять ДНК. З певним ймовірністю така куля може правильно передати генетичний матеріал клітині й рослина отримує нові властивості.
Перше повідомлення про успішне створення ГМ рослини з’явилося у 1983 році, де описувалось перенесення гену стійкості до комах у рослини тютюну. Першими ГМ рослинами, дозволеними для харчування людини, були FlavrSavr томати, створені каліфорнійською компанією Calgene. Ці томати мали покращену здатність до зберігання завдяки гену полігалактуронази. Вперше вони були комерціалізовані 1994 року в США. Станом на 2009 рік, через 15 років після початку комерціалізації, трансгенні рослини вирощувалися вже на 134 млн. га (9% від загальних 1,5 млрд. га світових площ сільськогосподарських земель). Зараз трансгенні рослини вирощують у 25 країнах, в яких проживає 3,6 млрд. або 54% світового населення. Шість країн з найбільшими площами вирощування генетично модифікованих культур – це США (64,0 млн. га), Бразилія (21,4 млн. га), Аргентина (21,3 млн. га), Індія (8,4 млн. га), Канада (8,2 млн. га), та Китай (3,7 млн. га). Решта 7 млн. га площ посівів трансгенних рослин припадають на 19 інших країн світу. З 1996 по 2009 рр. світові площі, на яких вирощуються ГМ рослини, зросли у 80 разів. Станом на 2009 рік ще у 32 країнах такі рослини були дозволені для ввезення та використання у якості продуктів харчування для людини і тварин.
8.Екологічна група рослин по відношенню до світла,фотоперіодизм.
По відношенню до світла всі рослини, в тому числі і лісові дерева поділяються такі екологічні групи:
· Геліофітів (світлолюбні), що потребують багато світла і здатні переносити лише незначне затінення (до світлолюбні ставляться майже всікактуси і інші сукуленти, багато представників тропічного походження, деякі субтропічні чагарники);
· Сціофіти (тіньолюбні) - задовольняються навпаки незначним освітленням і що можуть існувати в тіні (до тіньовитривалим відносяться різні хвойні рослини, багато папороті, деякі декоративно-листяні рослини);
· Тіньовитривалі (факультативні геліофітів).
Геліофітів. Світлові рослини. Мешканці відкритих місць проживання: лук, степів, верхніх ярусів лісів, ранньовесняні рослини, багато культурнірослини.
Характеризуються такими ознаками:
· Дрібні розміри листя; зустрічається сезонний диморфізм: навесні лестощі дрібні, влітку - більше;
· Листя розташовується під великим кутом, іноді майже вертикально;
· Листова пластинка блискуча або густо опушена;
· Утворюють виряджені насадження.
Сціофіти. Не виносять сильного світла. Місця проживання: нижні затемнені яруси; мешканці глибоких шарів водойм. Перш за все, це рослини, що ростуть під наметом лісу (кислиця, Костина, снить).
Характеризуються такими ознаками:
· Листи великі, ніжні;
· Листя темно-зеленого кольору;
· Листя рухливі;
· Характерна так звана листова мозаїка (тобто особливе розташування листя, при якому листя макимально не затуляють один одного).
Тіньовитривалі. Займають проміжне положення. Часто добре розвиваються в умовах нормального освітлення, але можуть при цьому переносити і затемнення. За своїми ознаками займають проміжне положення.
Причини цієї відмінності потрібно шукати, перш за все, в специфічних особливостях хлорофілу, потім в різній архітектоніці видів (в будові пагонів, розташуванні та формі листя). Розподіливши лісові дерева згідно з їх потребою в світі, що виявляється в їх змаганні, коли вони ростуть разом, і, ставлячи найбільш світлолюбні вперед, ми отримаємо приблизно наступні ряди.
1) Модрина, береза, осика, вільха.
2) Pinus silvestris, P. strobus, ясен, дуб, в'яз, Acer Pseudoplatanus.
3) Pinus montana Mill, ялина, липа, граб, бук, ялиця.
Чудово й біологічно важливо обставина, що майже всі дерева в молодості можуть переносити великого затінення е, ніж у більш зрілому віці. Далі слід зауважити, що здатність переносити затінення знаходиться у відомій залежності від родючості грунту.
Фотоперіодизм — фізіологічна реакція організмів на добовий ритм освітлення (співвідношення довжини дня та ночі). Зустрічається у рослин і тварин. Виявляється в коливаннях інтенсивності фізіологічних процесів. Найбільшою мірою фотоперіодизм властивий зеленим рослинам, життєдіяльність яких безпосередньо залежить від світлової енергії Сонця.
Фотоперіодизм відкрито в 1920 р. американськими вченими В. Гарнером і Г. Аллардом на прикладі тютюну.
Фотоперіодизм у рослин проявляється в зміні процесів росту та розвитку. Один з основних проявів цієї реакції – фотоперіодична індукція цвітіння.
За типами ФПР розрізняють такі основні групи рослин (Т. К. Горишина, 1980):
1. Рослини короткого дня, яким для переходу до цвітіння потрібно світла 12 і менше годин на добу (коноплі, тютюн);
2. Рослини довгого дня, для фази цвітіння їм потрібна довжина дня понад 12 годин на добу (картопля,пшениця, шпинат);
3. Рослини проміжного типу, цвітіння в яких наступає при певному діапазоні фотоперіодизму, наприкладгваюла;
4. Рослини фотоперіодично нейтральні, цвітіння в яких відбувається при будь-якій довжині дня (помідор,кульбаба тощо).
Особливо велику роль фотоперіодизм відіграє в географічному поширенні рослин і в регуляції їх сезонного ритму.
Фотоперіодизм вивчали американські фізіологи рослин У. Гарнер, Г. Аллард у 1920-1925 рр. В залежності від реакції рослин на довжину дня, що прискорює зацвітання, вони поділяються на:
· довгоденні (ДДР - рослини довгого дня – злаки, хрестоцвіті, кріп; поширені в основному в помірних та приполярних широтах);
· короткоденні (КДР - рис, соя, конопля; субтропіки);
· довгокороткоденні (ДКДР) і
· короткодовгоденні (КДДР) – потребують чергування різних фотоперіодів;
· нейтральні (НДР - гречка, горох і ін.).
Поділ рослин на дані групи не пов’язаний з конкретною оптимальною довжиною дня, а лише дає уявлення про те, чи прискорюється цвітіння при збільшенні чи зменшенні тривалості освітлення в кожному фотоперіоді.
Є рослини з якісними й кількісними типами фотоперіодичних реакцій. Інтенсивність освітлення, температура й інші фактори можуть змінювати характер фотоперіодичних проявів рослин.
Для КДР із якісною реакцією основним фактором служить тривалість темнового періоду. Його переривання (хоч на 1 хв.) перешкоджає переходу до цвітіння. Переривання світлового періоду темнотою не впливає на строки цвітіння. ДДР не потребують періоду темноти і зацвітають при безперервному освітленні.
Фотоперіодична дія сприймається головним чином листками, а не апексами пагонів. Максимальну чутливість до фотоперіоду мають листки, які тільки що закінчили ріст. Здійснюється це головним чином завдяки фітохрому. Вважають, що його темнове перетворення може служити способом вимірювання часу по типу піскового годинника. Спалах червоного світла встановлює фазу ендогенного ритму.
Необхідність великої поверхні листків і достатньої інтенсивності світла для переходу до цвітіння у багатьох рослин пояснюється потребою ростучих меристем в асимілятах. У НДР перехід до цвітіння забезпечується віковими змінами. Тут спрацьовує ендогенна регуляція. Дані фактори (температура і фотоперіод) можуть діяти послідовно, як, наприклад, в озимих злаків.
Температурна й фотоперіодична регуляція служать пристосуванням рослин до умов існування, тому що обумовлюють сприятливі строки для переходу до цвітіння. В ході фотоперіодичної індукції в листках утворюється стимулятор цвітіння, який транспортується у вегетативні бруньки пагонів, де включає другу фазу ініціації – евокацію.
7. Транспорт сполук по клітиніМЕМБРÁННИЙ ТРÁНСПОРТ (лат. transporto — переношу, перевожу, переміщаю) — перенесення різноманітних речовин через плазматичну мембрану (див. Мембрана біологічна). Механізм транспорту речовин до клітини та з неї залежить від розмірів частинок, що транспортуються. Малі молекули та іони проходять крізь мембрани шляхом пасивного чи активного транспорту. Перенесення макромолекул та великих частинок здійснюється за рахунок ендоцитозу та екзоцитозу.
Пасивний транспорт відбувається мимовільно без затрати енергії шляхом дифузії, осмосу та полегшеної дифузії. Дифузія — це транспорт молекул та іонів через мембрану з ділянки з високою концентрацією до ділянки з низькою концентрацією, тобто речовини надходять за градієнтом концентрації. Дифузія може бути простою та полегшеною. Якщо речовини добре розчинні в ліпідах, то вони проникають до клітини шляхом простої дифузії. Напр. оксиген, потрібний клітині при диханні, та вуглекислий газ у розчині швидко дифундують крізь мембрани. Таким способом проникають до клітини також деякі фармацевтичні препарати, які є ліпідорозчинними. Вода також здатна проходити крізь мембранні пори, що утворені білками, і переносити молекули та іони речовин, які в ній розчинені. Дифузію води крізь напівпроникну мембрану називають осмосом. Вода переходить з ділянки з низькою концентрацією солей до ділянки, де їхня концентрація вища. Тиск на мембрану, що виникає при цьому, називають осмотичним. Усі живі клітини здатні регулювати осмотичний тиск, змінюючи концентрацію речовин поза клітиною та всередині клітини. Речовини, які не є розчинними у ліпідах, транспортуються через іонні канали, утворені в мембрані білками, чи за допомогою білків-переносників. Це полегшена дифузія, шляхом якої, напр. здійснюється надходження глюкози до еритроцитів. Серед систем пасивного транспорту важливу роль відіграють іонні канали, які забезпечують проникність мембрани для Na+, K+, Ca2+. Na+-канали активуються вератрадином, батрахотоксином, блокуються амілоридом, тріамтереном; K+-канали блокуються місцевими анестетиками (лідокаїном, дикаїном), деякими протисудомними (дифеніл, карбамазепін, вальпроати, фенобарбітал та ін.) і протиаритмічними засобами (аміодарон); Ca2+-канали чутливі до цілої низки хімічних речовин, зокрема верапамілу, дилтіазему, ніфедипіну та інших похідних дигідропіридинів. Трансмембранний обмін (антипорт) чи односпрямований транспорт (симпорт) іонів здійснюється спеціальними білками-переносниками. Система односпрямованого транспорту (котранспорту) представлена (Na++K++Cl–)-переносником, що є чутливим до дії діуретиків (фуросеміду, амілориду, туметаніду). Виключення або різка зміна властивостей переносників і каналів лежить в основі дії багатьох токсичних і фармацевтичних речовин. Деякі речовини — іонофори, до яких належать різні антибіотики (валіноміцин, амфотерицин В, нонактин, енніатини, аламетицин та ін.), синтетичні циклополіефіри самостійно здатні утворювати канали у ліпідному бішарі мембрани. Дія деяких ЛП заснована на зміні властивостей каналів і переносників, що дозволяє регулювати транспорт речовин у клітинах і організмі в цілому.
Активний транспорт речовин крізь мембрану здійснюється проти градієнта їхньої концентрації із затратою енергії АТФ та за участю спеціальних мембранних білків — транспортних АТФаз, які також називаються іонними насосами. Найбільш поширеними в клітині тварин є Н+-АТФаза, Na+,K+-АТФаза і Са2+-АТФаза, що являють собою цілі мембранні комплекси із складною структурою. Функціональне значення біологічних насосів полягає у підтримці всередині клітини постійного іонного складу. Na+,K+-АТФаза сприяє виведенню Na+ з клітини та надходженню К+ до клітини за допомогою енергії АТФ і є прикладом антипортного транспорту. Із впливом на натрієвий насос пов’язаний механізм дії деяких фармацевтичних препаратів. Так, напр. серцеві глікозиди (дигоксин, уабаїн, строфантин К) пригнічують Na+,K+-АТФазу; деякі діуретики (тіазиди) інгібують активний транспорт Na+ та/або Cl– в епітелії канальців нирок; омепразол знижує кислотність шлункового соку, незворотно пригнічуючи протонний насос Н+-АТФазу парієтальних клітин шлунка. Са2+-АТФаза високочутлива до дії різноманітних тіолових отрут та ін. Крім вищенаведених видів активного транспорту, виділяють специфічні механізми переміщення речовин, пов’язані з порушенням цілісності мембрани, ендоцитоз та екзоцитоз. При ендоцитозі плазматична мембрана утворює вирости, які потім перетворюються на внутрішньоклітинні пухирці, що містять захоплений клітиною матеріал. Ці процеси відбуваються із витратою енергії АТФ. Розрізняють два види ендоцитозу: фагоцитоз і піноцитоз. Фагоцитоз (грец. phagos — пожирати, cytos — клітина) — це захоплення і поглинання клітиною великих часток (іноді цілих клітин та їхніх частин). Ендоцитоз рідини та розчинених в ній речовин називається піноцитозом (грец. pyno — пити, cytos — клітина). Шляхом ендоцитозу, напр. відбувається всмоктування жиру клітинами кишкового епітелію. Екзоцитоз — це процес виведення з клітини різноманітних речовин крізь мембрану, фактично зворотний ендоцитозу механізм. Шляхом екзоцитозу вивільнюються гормони, жирові краплини, а також медіатори в синапсах при збудженні.
Таким чином, знання про особливості М.т. є дуже важливими для фармації, оскільки визначають не лише шляхи проникнення всіх фармацевтичних препаратів до певних клітин, але й безпосередню дію багатьох з них.
6.Метоз,мійоз,сигнальні закінчення білків.
Мітоз (рідше: каріокінез або непрямий поділ) (від грец. mitos — нитка) — поділ ядра [етап кон'югаціюгомологічних хромосом протягом профази.
Історія
У 1874І. Д. Чистяков описав ряд стадій (фаз) мітозу у спорах плаунів, ще не ясно представляючи собі їхню послідовність. Детальні дослідження з морфології мітозу уперше були виконані Е. Страсбургером на рослинах (1876 — 1879) і В. Флеммінгом на тварин (1882).
Фази мітозу
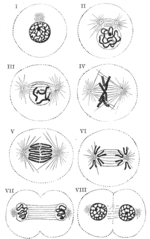

Мітоз. I — III — профаза; IV — метафаза; V — VI — анафаза; VII — VIII — телофаза.
Мітоз — лише одна з частин клітинного циклу, але він достатньо складний, щоб в його складі, у свою чергу, було виділено чотири фази: профаза, метафаза, анафаза і телофаза. Подвоєння хромосом відбувається ще в ході інтерфази. В результаті цього, в мітоз хромосоми вступають вже подвоєними, такими, що нагадують букву «X» (ідентичні копії материнської хромосоми сполучені один з одним в області центромери).
У профазі відбувається конденсація хромосом, клітинний центр ділиться і продукти його поділу розходяться до полюсів ядра, руйнується ядерна оболонка, утворюється веретено поділу.
У метафазі хромосоми розташовуються на «екваторі» (на рівній відстані від «полюсів» ядра) в одній площині, утворюючи так звану метафазну пластинку. Важливо відзначити, що вони залишаються в такому положенні протягом досить тривалого часу. Зазвичай у зв'язку з цим метафаза — найслушніший час для підрахунку хромосомних чисел.
У анафазі, яка є найкоротшою фазою мітозу, хромосоми діляться (з'єднання в районі центромери руйнується) і розходяться до різних полюсів клітини.
У телофазі відбувається руйнування веретена поділу і утворення ядерної оболонки навколо дочірніх ядер.
Тривалість мітозу в середньому 1-2 год., різна для різних видів клітин. Процес залежить також і від умов зовнішнього середовища (температури, світлового режиму й інших показників).
Варіанти мітозу
Слід зазначити, що в різних групах живих організмів мітоз протікає дещо по-різному. Описаний вище варіант мітозу називається відкритий ортомітоз (ядерна оболонка руйнується, веретено поділу пряме, оскільки продукти поділу клітинного центру розміщені на протилежних полюсах ядра). Характерний для багатоклітинних тварин, багатоклітинних рослин і ряду найпростіших.
У деяких групах найпростіших продукти поділу клітинного центру в анафазі не досягають протилежних сторін ядра, внаслідок чого мікротрубочки веретена поділу розташовуються під кутом, нагадуючи букву V (такий варіант ділення отримав назву плевромітоз). У ряді випадків мітоз відбувається без руйнування ядерної оболонки (закритий мітоз). Нарешті, клітинний центр може містити центріолі (наприклад, у тварин) або не містити їх (наприклад, у квіткових рослин). Відповідно, розрізняють також центріолярний і ацентріолярний мітоз.
Біологічне значення мітозу
Біологічне значення мітозу полягає в строго однаковому розподілі редуплікованих хромосом між дочірними клітинами, що забезпечує утворення генетично рівноцінних клітин і зберігає наступність у ряді клітинних поколінь.
Мейоз (або редукційний поділ) — особливий вид поділуеукаріотичнихклітин, характерний тільки статевим клітинам (не соматичним), унаслідок якого хромосомний набір зменшується вдвічі, клітини переходять з диплоїдного стану в гаплоїдний.
Мейоз складається з двох послідовних поділів, аналогічних мітотичним (з деякими відмінностями), інтерфаза між якими вкорочена, а у рослинних клітинах може бути взагалі відсутня.
| 1 Історія вивчення мейозу 2 Процес мейозу 2.1 Інтерфаза-І 2.2 Профаза-І 2.3 Метафаза-І 2.4 Анафаза-І 2.5 Телофаза-І 2.6 Інтерфаза-ІІ 2.7 Профаза-ІІ 2.8 Метафаза-ІІ 2.9 Анафаза-ІІ 2.10 Телофаза-ІІ 3 Біологічне значення мейозу 4 Посилання |
Історія вивчення мейозу
Мейоз був вперше вивчений і описаний у яйцях морських їжаків німецьким біологомОскаром Гертрігом у 1876 році.
У 1883 році мейоз був знову описаний, уже на хромосомному рівні, бельгійським вченим Едуардом фон Бенеденом.
Проте важливість мейозу у спадковості була описана лише у 1890 році німецьким біологом Августом Вайсманом.
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 2014; Нарушение авторских прав?; Мы поможем в написании вашей работы!