КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биологическое окисление. Биоэнергетика
|
|
|
|
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 25
по дисциплине «Методы подсчета запасов и ресурсов»
1. Подсчёт запасов категории С1 – какие объекты подлежат такому обсчёту.
2. Основные промысловые параметры продуктивных горизонтов.
3. Случаи, когда пересчитывают и переутверждают запасы в ГПЗ РФ.
Экзаменационные билеты рассмотрены на заседании кафедры №___ от______________2012 г.
Зав. кафедрой, профессор _______________________________ С.П. Примина
Жизнь - совокупность химических реакций (эндоэргических и экзоэргических). Одно из свойств живой материи - способность извлекать из окружающей среды энергию для поддержания внутреннего постоянства (2-е начало термодинамики).
по способу
/ \
аутотрофы гетеротрофы (энергия
(фотосинтез) химических связей)
Энергия идет на обеспечение различных видов биологической работы: реакции синтеза, механическое движение, осмос, перенос через мембрану, передача нерв-
ных импульсов. Способ извлечения энергии химических связей - окислительно - восстановительные реакции (биоокисление).
Раздел биохимии, занимающийся вопросами преобразования и использования энергии в живых клетках, называется б и о э н е р г е т и к о й.
2 вида полезной энергии
/ \
свободная тепловая
(производит работу при const и Р) (производит работу при > t, Р)
Сходство процессов биоокисления и горения:
1. Конечные продукты СО2 и Н2О - одинаковы;
2. Выделяемое количество энергии - одинаково;
3. Окисляются одни и те же субстраты.
Различия:
1. Горение - одноэтапный процесс (присоединение О2 к веществу. Биоокисление - многоэтапный);
2. При горении высвобождается все количество энергии сразу, при биоокислении - постепенно в виде небольших порций и на отдельных этапах идет аккумулирование энергии;
3. При горении не нужны ферменты, при биоокислении присутствуют ферменты;
4. При горении основная масса энергии выделяется в результате окисления С С + О2 - СО2, при биоокислении идет процесс дегидрирования. 2Н + 1/2 О2 - Н2О.
Химические реакции, в процессе которых происходит перенос электронов от од- ной молекулы к другой называется окислительно - восстановительными реак- циями. Соединения, отдающие электроны в такой реакции, называются донорами электронов или восстановителями, а соединения, присоединяющие электроны - акцепторами электронов или окислителями. Окислители и восстановители всег- да реагируют как сопряженные окислительно - восстановительные пары.
Способность каждой окислительно - восстановительной пары - обратимо отда- вать электрон - выражают стандартным окислительно - восстановительным потенциалом.Ео `. В биохимии для выражения стандартных потенциалов принято пользоваться понятием восстановительный потенциал. Чем более отрицатель- ной величиной выражается восстановительный потенциал системы, тем выше ее способность отдавать электроны; и наоборот, чем более положительный восста- новительный потенциал, тем выше способность системы присоединять электроны.
В эукариотических клетках процесс биологического окисления происходит во внутренней мембране митохондрий, где локализуются переносчики электронов, составляющие дыхательную цепь и органы, катализирующие синтез АТФ из АДФ и фосфата.
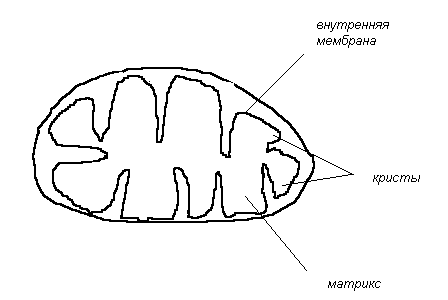
Н а р у ж н а я м е м б р а н а - легко проницаема для всех молекул и ионов не- большого размера.
В н у т р е н н я я м е м б р а н а - содержит цепь переноса электронов, АТФ - син- тетазу и различные транспортные системы, для большинства ионов небольшого размера она непроницаема.
М а т р и к с - содержит фрагменты ЦТК, систему окисления жирных кислот и др.
К р и с т ы - выпячивания внутренней мембраны.
Таким образом, внутренняя мембрана митохондрий состоит из множества ком- понентов. В нее входят переносчики электронов, ряд ферментов и некоторые транспортные системы. На долю этих компонентов приходится в общей сложности до 75 % или даже больше общей массы мембраны. Внутренняя митохондриальная мембрана имеет сложную мозаичную структуру, от целостности которой зависит такая жизненно важная функция, как синтез АТФ.
Б и о л о г и ч е с к о е о к и с л е н и е - это совокупность окислительно - восстановительных реакций, совершающихся в биологических объектах под влиянием ферментов.
Биологическое окисление, в котором конечным акцептором электрона и Н явля- ется кислород, называется тканевым дыханием (это основной процесс, который позволяет запасать энергию впрок в больших колличествах).
Если же конечным акцептором электрона в процессах биологического окисления будет не кислород, а какое - либо другое соединение - это субстратное окисление - это окисление в анаэробных условиях, дает малый выход энергии, но это единственная возможность энергию клеткой в условиях гипоксии.
Окисление субстратов в процессе дыхания можно представить как перенос электронов и ионов Н от органических веществ на кислород. Этот процесс включает много этапов, в нем учавствуют ряд промежуточных переносчиков, образующих цепь переноса электронов и протонов или дыхательную цепь. Промежуточными акцепторами электронов и ионов Н при биоокислении являются окислительно-восстановительные ферменты (класс оксидоредуктаз). Особенность этих ферментов то, что они работают в виде мультиферментных систем (дыхательная цепь, дыхательные ансамбли), потому что образуют с клеточной структурой надиолекулярные комплексы (работают и входят в состав клеточных мембран (внутри кристы митохондрий расположен в строго определенной последовательности, этим обеспечивается высокая скорость и эффективность процесса, исключающая к минимуму побочные реакции.
Электроны и протоны для большинства субстратов поступают на НАД или НАДФ - пиридинзависимые дегидрогеназы. Они катализируют обратимые реакции, кото- рые в общем виде могут быть записаны так: +
Восстановленный субстрат + НАД = окислительный субстрат + НАДН + Н
Большинство таких дегидрогеназ содержат НАД (активная форма Vif В5) у неко- торых, как например глюкоза 6-фосфатдегидрогеназы, акцептором является НАДФ.
Основными о с о б е н н о с т я м и такого переноса 2Н и електронов являются:
а) обратимость многих дегидрогеназных реакций вследствие этого, что измене- ние энергии при этих реакциях как правило невелико (участие не только в процес- се распада, но и синтеза веществ);
б) способность коферментов (как в окисленной,так и в восстановленной форме -легко отделяться путем диссоциации от апофермента (обмен между различными дегидрогеназами, синтез веществ);
в) возможность участия в таких реакциях субстратов различных типов - углевод, липид, аминокислот и т.д.).
Пиридинзависимые дегидрогеназы отщепляют от своих S по 2Н и 2 электрона, один Н в виде гидрид - иона переносится на НАД или НАДФ, а второй в виде Н пе- ходит в среду, каждый гидрид - ион несет 2 восстановительных эквивалента; один из них в форме водородного атома присоединяется к четвертому углеродному атому никотинамидного кольца, а другой в виде электрона передается N этого кольца:
Поскольку большая часть клеточных дегидрогеназ переносят водородные ато- мы от субстратов, можно сказать, что НАД выполняет коллекторную функцию - со- бирает электроны, поступающие от разных субстратов в одной молекулярной фор- ме НАДН.
Следующим переносчиком, принимающим электроны и Н от НАДН являются флавопротеиды (восстановленная форма - желтая) содержит кофермент ФАД и и ФМН - витамин В2 (рибофлавин, способный присоединять 2 электрона и 2Н. От- личием от НАД - прочная связь с апоферментом:
Реакции, катализируемые ФАД - зависимыми дегидрогеназами легкообратимы. Флавопротеиды функционируют в нескольких пунктах, где электроны включаются в дыхательную цепь.
Окисление некоторых субстратов начинается с ФАД - зависимых дегидрогеназ (сукцинат, жирные кислоты).
Роль следующего звена играет убикинон или кофермент Q (производное Vit h):
Молекулы убикинона выполняют коллекторную функцию, собирая восстановлен- ные эквиваленты не только от НАДН - дегидрогеназ, но и от других флавиновых дегидрогеназ. Кроме того, его небольшой удельный вес и растворимость в липи- дах митохондриальной мембраны идеально соответствует функции Ко Q, как под- вижного переносчика, взаимодействующего с фиксированными электронперенося- щими белками.
Следующим компонентом дыхательной цепи, принимающим электроны от убики- нона, являются цитохромы простетической гр.-гем. Протоны уходят в среду. Су- ществует 3 класса цитохромов: а, б и с, различающихся по спектрам поглощения. Цитохромы в дыхательной цепи располагаются в следующей последовательности в - С1 - С - аа3.
Цитохром в принимает электроны от убихинона и передает их на цитохром С1, кото-рый в свою очередь передает их на цитохром С. Каждый из этих цитохромов, нахо-дясь в окисной Fe (III) форме, присоединяет один электрон и переходит в закисную Fe (II) форму. В переносе электронов от убихинона на цитохром С принимает учас- тие так же белок, содержащий железо и серу. Последний в ряду переносчиков элек- тронов стоит цитохром аа3, называемый также цитохромоксидазой, он переносит электрон прямо на О2, и тем самым завершает процесс переноса электронов. Этот этап является очень важным, т.к. должны на кислород быть переданы 4 элек-трона, для того чтобы образовались две молекулы воды (четыре Н-иона, которые тоже для этого необходимы, поступают из водной среды). Из всех переносчиков цепи переноса электронов только цитохром аа3 способен непосредственно всту- пать непосредственно в реакцию с кислородом. Для клетки это очень важно, пото- му что при неполном восстановлении кислорода в случае присоединения только 2-х электронов, образуется перекись водорода (Н2О2), а в случае присоединения од- ного электрона - супероксидный радикал (:О2). И перекись, и супероксид крайне токсичны для клеток, потому что они повреждают клеточные мембраны, взаимо-действуя с остатками ненасыщенных жирных кислот мембранных липидов. Аэроб-ные клетки защищают себя от этого вредного действия супероксида и перекиси с помощью двух ферментов: супероксиддисмутазы (превращающего супероксид- ный радикал в перекись водорода) и каталазы (превращающий Н2О2 в Н2О и 1/2 О2:
Электроны переходят от электроотрицательных систем к электроположительным связям с тем, что такой поток сопровождается уменьшением свободной энергии; поток электронов направлен так, чтобы в результате свободная энергия системы уменьшалась. Чем больше разность стандартных потенциалов двух окислительно - восстановительных пар, тем больше уменьшение свободной энергии при переносе электронов от электроотрицательной пары к электроположительной. Пройдя через всю цепь переносчиков электронов от НАД (- 0,326) к кислороду (+ 0,826) элект- роны теряют значительное количество свободной энергии, поскольку разница меж- ду стандартными потенциалами пар НАД/ НАДН и Н2О/ 1/2 О2 относительно вели- чин 1,146 = 52,6 ккал. Этой энергии более чем достаточно для синтеза 3 молекул АТФ, которые в стандартных условиях требуют затраты 3х7,5 = 22,5 ккал (40%).
Наши данные свидетельствуют о том, что электроны движутся в сторону кисло- рода:
во-первых, именно в это последовательности их ред. окспотенциалы становятся все более положительны по мере приближения к кислороду, а этого и следует ожи- дать, поскольку электроны стремятся переходить от электроотрицательного к электроположительному, что вызывает снижение свободной энергии;
во-вторых, каждое звено этой цепи специфично в отношении определенного до- нора и определенного акцептора электронов;
в-третьих, из митохондриальной мембраны удалось выделить структурно обо- собленные комплексы функционально связаных между собой переносчиков элект- ронов.
Около 40% энергии запасается в виде АТФ, а остальная энергия выделяется в виде тепла, являющегося одинаково важнейшим фактором в работе механизмов терморегуляции, который поддерживает изотермическое состояние клетки нашего организма могут использовать энергию химических связей.
При переносе электронов и протонов по дыхательной цепи происходит выделе- ние энергии на каждом этапе, где перепад ред-окспотенциалов большой, происхо- дит выделение энергии, достаточное для синтеза АТФ. Всего таких точек 3:
1) между НАД/ФАД
2) цит. в/с
3) цит. аа3/ 1/2 О2, т.е. максимально при окислении с НАД - 3 АТФ.
с ФАД - 2 АТФ
Следовательно, дыхательная цепь представляет собой своего рода каскад, при помощи которого клетка получает свободную энергию, извлекаемую из клеточного топлива, в "расфасованном" и следовательно удобном для использования виде.
7,5 х 3 = 22,5 ккал (40%)
Изучение о переносе электронов в немалой мере способствовал и такой метод, как применение ингибиторов, блокирующих определенные этапы этапы этого про- цесса.
1) Ротенон аминобарбитал блокирует перенос электронов на участке от НАДН до ФАД (это высокотоксичное соединение, добываемое из растений, употреблялось индейцами в качестве яда для рыб);
2) Токсичный антибиотик антимицин А, блокирует перенос электронов от убихи- нона на цитохром С;
3) Цианид - один из самых сильных ядов, блокирующий процесс восстановления кислорода, катализируемый цитохромоксидазой аа3 (СО - окись углерода).
Окислительное фосфорилирование - это синтез АТФ из АДФ и Фн в процессе тка- невого дыхания.
Субстратное фосфорилирование - процесс переноса макроэргической связи помимо дыхательной цепи за счет окисления, либо за счет молекулярной перестр- ройке субстрата, т.е. без участия кислорода в анаэробных условиях.
Об эффективности процессов окислительного фосфорилирования судят по ве- личине отношения этерифицированного фосфата к поглощенному кислороду (О), т.е. по отношению р/о
3 АТФ - р/о = 3
2 АТФ - р/о = 2
1 АТФ - р/о = 1
Сопряженные реакции
Пример: Глю + Н3РО4 -
Окислительное фосфорилирование - синтез АТФ из АДФ и Фн в процессе тканево- го дыхания.
Химиосмотическая теория Митчелла (1961) и далее была развита Скулачевым.
Скорость окислительного фосфорилирования в первую очередь зависит от со- держания АДФ в клетке, чем быстрее расходуется АТФ при выполнении свойствен- ных организму работ, тем больше накапливается АДФ, тем больше потребность в энергии и следовательно в синтезе АТФ.
Регуляция скорости окислительного фосфорилирования содержанием АТФ называют дыхательным контролем.
Макроэргическая связь - связь, богатая энергией - это химическая связь, при разрыве которой выделяется 5 и более ккал/моль энергии. Соединения, содержа- щие эту связь, называются макроэргическими. Обозначают ее (~) извилистой ли- нией. При разрыве одной связи АТФ выделяется 7,3 ккал/моль. Для синтеза одной макроэргической связи АТФ требуется столько же энергии. Следовательно, при переносе пары атомов водорода и пары электронов по дыхательной цепи синтези- руется и запасается 7,3 ккал/мольх3 = 21,9 ккал/моль, это составляет 40% всей высвободившейся при переносе энергии АТФ - важнейший макроэрг, используе- мый в организме, но не единственный. Все макроэргические соединения можно разделить на 3 группы:
1) высокоэргические - ФЭП - 14,8 ккал/моль, 1,3 ДФГ - 11,8 ккал/моль, КФ - 10,3 ккал/моль;
2) низкоэргические - глю - в ф - 3,3 ккал/моль, глю/ф - 5,0 ккал/моль, фру 6 - ф - 3,8 ккал/моль;
3) АТФ - 7,3 ккал/моль.
АТФ функционирует в клетке как общий промежуточный продукт, переносящий энергию от реакций, сопровождающиеся выделением свободной энергии, с теми, в которых потребляется энергия. Протекающие ферментативные реакции в клетке переноса фосфатных групп, важнейшая особенность этого переноса состоит в том, что почти все высокоэргические фосфорные соединения переносят свои группы низкоэргическим акцепторам фосфата через АТФ с помощью специфичес- ких киназ.
Использование энергии АТФ в организме в процессе жизнедеятельности в виде 4-х видов работ: 1) механическая
2) осмотическая
3) химическая
4) энергетическая.
|
|
|
|
|
Дата добавления: 2015-05-08; Просмотров: 2276; Нарушение авторских прав?; Мы поможем в написании вашей работы!