КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транскрипционных факторов
|
|
|
|
При научении в мозге активируются гены
Генов в мозге
Формирование нового опыта требует экспрессии
Современные представления о молекулярно-биологических механизмах обучения и памяти строятся на положении о кратковременной и долговременной формах хранения информации в мозге (гл. 6). В основе этой теории лежит открытие Г. Мюллера и А. Пильзекера, обнаруживших в 1900 г., что переход из кратковременной и легко нарушаемой памяти в долговременную и устойчивую память происходит у человек в течение первого часа после получения им новой информации. Они назвали этот процесс консолидацией памяти (Muller, Pilzecker, 1900).
Основным шагом в понимании биологических механизмов консолидации памяти стало открытие 1960-х гг., показавшее, что переход памяти из кратковременной в долговременную форму требует синтеза новых молекул РНК и белка, т.е. экспрессии генов. Было установлено, что волна синтеза новых белков в клетках при запоминании информации совпадает с периодом консолидации памяти, обнаруженным Мюллером и Пильзекером, а химическая блокада экспрессии генов в этот период нарушает образование долговременной памяти (Davis, Squire, 1984). Оказалось также, что «критическое окно» амнестического действия блокаторов экспрессии генов универсально для самых разных видов обучения и различных организмов, от беспозвоночных до человека (Goelet et al., 1986). Данное предположение также хорошо согласовывалось с гипотезой об участии клеточного роста и изменения морфологии синапсов в долговременной памяти (Tanzi, 1893; Hebb, 1949).
Таким образом, понятие долговременной памяти постепенно трансформировалось из условного обозначения относительной продолжительности явления в компонент биологической концепции, связывающей научение и опыт с морфогенезом и развитием (Bailey, Kandel, 1994). Критическим звеном этой концепции стал молекулярный механизм консолидации памяти, отождествляемый с активацией транскрипции генов в нервных клетках при научении. Однако то, какие именно гены активируются в при научении и каковы их функции в нервных клетках, долгое время оставалось неизвестным.
Первыми генами, активация которых была обнаружена в мозге при обучении, оказались так называемые «непосредственные ранние гены», кодирующие транскрипционные факторы.
«Непосредственные ранние гены» были впервые обнаружены при изучении механизмов геномного ответа на действие факторов роста, запускающих процессы клеточного цикла (Greenberg, Ziff, 1984; Lau, Nathans, 1985). Индукция их транскрипции происходила несмотря на подведение ингибиторов синтеза белка, то есть строилась на механизмах, заранее готовых для восприятия экстраклеточных стимулов. Первые из идентифицированных продуктов генов данного семейства оказались ядерными белками, связывающимися с ДНК и регулирующими транскрипцию других генов. По этим свойствам данные гены значительно напоминали группу «непосредственных ранних генов» бактериофагов и эукариотических ДНК-вирусов, поэтому, по аналогии с вирусными генами, эта группа быстро активирующихся генов получила название «клеточных непосредственных ранних генов» (Lau, Nathans, 1985). Это же семейство часто обозначается как «гены первичного ответа», «гены раннего ответа» или просто «ранние» гены (Анохин, 1997).
Одним из первых в данной группе был клонирован ген c-fos. Его структура и свойства хорошо изучены, и он может служить прототипом генов данного семейства. Первоначально было установлено, что в ходе эмбрионального развития c-fos играет важную роль в регуляции процессов клеточного роста и пролиферации. Оказалось также, что один белок, кодируемый геном c-fos, сам по себе не может инициировать или подавить транскрипцию генов-мишеней, необходимых для инициации клеточного деления или дифференцировки. Он должен образовать димеры с молекулами других транскрипционных факторов, объединяемых в семейство, получившее название АР-1 (Curran, Morgan, 1987). Помимо c-fos в это семейство входят fos-B, c-jun, jun-B,jun-D,fra- 1,fra-2 и ряд пока еще не идентифицированных генов. Кроме того, были клонированы и другие ранние гены, такие как ets-1, ets-2, Мус, Myb, Krox-20, zif/286, NGFI-B, mKr2, Arg3.1 (Sheng, Greenberg, 1990; Herdegen, Leath, 1998). Продукты многих из них, хотя и не всех, также являются регуляторными белками, контролирующими транскрипцию. Гены, экспрессия которых находится под контролем индуцируемых транскрипционных факторов, были названы, по аналогии с вирусными системами, «поздними» генами, «генами позднего ответа» или «эффекторными» генами (Curran, Morgan, 1987), а весь двухфазный механизм регуляции транскрипции с участием этих двух классов генов является одним из наиболее универсальных способов обеспечения процессов клеточного деления и роста в развитии (рис. 20.4).

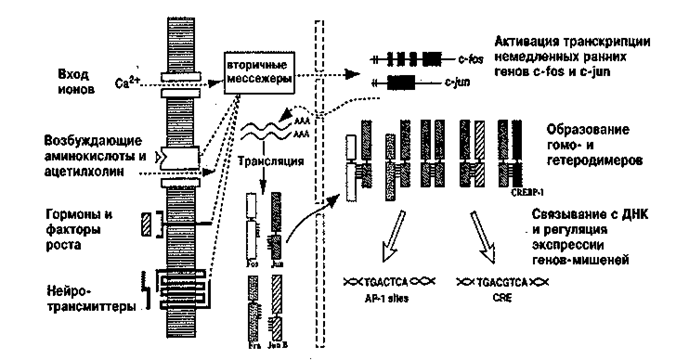
Рис. 20.4. Двухфазная регуляция транскрипции генов в клетке с
помощью продуктов «ранних» генов
Внеклеточные стимулы (гормоны, факторы роста, нейромедиаторы) могут вызвать активацию транскрипции «ранних» генов, включая гены c-fos и c-jun. Воздействие экстраклеточных сигналов на промоторы этих генов осуществляется посредством вторичных мессенджеров. Fos,Jun и ряд других белков этого семейства синтезируются в цитоплазме и быстро траспортируются в ядро, где могут образовывать гетеро- и гомодимерные комплексы. Эти комплексы обладают специфической ДНК-связывающей активностью и способны изменять транскрипцию других генов-мишеней («поздних» генов)
В середине 1980-х гг. несколько исследовательских групп обнаружили экспрессию гена с-fos в мозге обучающихся взрослых животных. Это повлекло за собой широкий спектр исследований участия этого и других «непосредственных ранних генов» в самых разных задачах обучения. Некоторые из результатов этих исследований суммированы в таблице 20.2.
Таблица 20.2
Особенности экспрессии непосредственных ранних генов (НРГ)
|
|
|
|
|
Дата добавления: 2015-05-28; Просмотров: 484; Нарушение авторских прав?; Мы поможем в написании вашей работы!