КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Движение прокариотических клеток
|
|
|
|
Биофизика сократительных систем. Молекулярные основы биологической подвижности. МОЛЕКУЛЯРНЫЕ МОТОРЫ.
Основные типы сократительных и подвижных систем. Немышечные формы подвижности клеток. Молекулярные механизмы немышечной подвижности. Структура и функционирование поперечнополосатой мышцы позвоночных. Молекулярные механизмы подвижности белковых компонентов сократительного аппарата мышц. Принципы преобразования энергии в механохимических системах.
Движение одноклеточных и многоклеточных организмов в пространстве, движение органоидов и других компонентов внутри клетки осуществляется за счет работы специфических механохимических преобразователей энергии, т.н. молекулярных моторов. В настоящее время известны два типа таких двигателей, использующих энергию различных источников: потребляющие энергию переноса ионов и потребляющие энергию АТФ. К первому типу относятся флагеллярные моторы, которые обеспечивают движение бактерий и протонные АТР-синтазы. Оба эти двигателя содержат вращающиеся детали и движущей силой для их функционирования является энергия разности концентрации ионов водорода на мембране (электрохимический потенциал). Ко второму типу молекулярных моторов относятся белки актомиозинового комплекса мышечных клеток, комплекс двигательных белков одноклеточных эукариот, белок кинезин и другие двигательные белки, выполняющие функции переносчиков органоидов (митохондрий, лизосом и др.). Ниже будут рассмотрены механизмы генерации и реализации движения у прокариот, у одноклеточных эукариот, у многоклеточных организмов. Также будут приведены сведения о механизме осуществления движения внутриклеточных структур.
Движение бактерий в пространстве происходит за счет вращения специальных образований на мембране - жгутиков. Жгутик представляет собой нитевидный вырост с поверхности бактерии. Жгутик состоит филамента, крюка и базального тела. Филамент образован полимерным белком флагеллином. Мономером флагеллина является глобулярный белок с молекулярной массой около 53 кД и диаметорм глобулы около 4,5 нм. Филамент состоит из нескольких флагеллиновых тяжей, закрученных в левую спираль. Крюк жгутика служит связкой между филаментом и базальным телом. Он состоит из белковых глобул. Базальное тело представляет собой сложную структуру, ответственную за генерацию движения. Он представляет собой комплекс белковых молекул и мембранных липидов (Рис. 1). Движение бактериальной клетки осуществляется за счет быстрого вращения жгутика (жгутиков). Электромоторы бактерий работают очень эффективно и экономично. Скорость вращения жгутика доходит до 50-100 оборотов в секунду (у некоторых видов бактерий скорость вращения превышает 1000 об/с.). При этом на генерацию движения расходуется не более 1% энергетических ресурсов бактериальной клетки. В жидкой среде бактерии двигаются со средней скоростью около 25 мкм/с, а некоторые виды могут двигаться поступательно со скоростью больше 100 мкм/с.
Механизмы генерации движения бактериальной клетки рассмотрим на примере Е. сoli. Из тела бактерии наружу выступают шесть жгутиков, каждый из которых представляет собой спиралевидную нить диаметром 15 нм и длиной 10 мкм. Когда жгутики начинают синхронно вращаться против часовой стрелки, они сплетаются в единый пучок, который образует своеобразный пропеллер (рис 1, А). Вращение пропеллера создает силу, заставляющую бактерию двигаться почти по прямой линии. Для прекращения движения в данном направлении, жгутики начинают вращаться в противоположную сторону, пучок расплетается, и бактерия останавливается. Она начинает хаотически вращаться, ее ориентация изменяется. В тот момент, когда все жгутики бактерии снова начнут синхронно вращаться против часовой стрелки, образовав пропеллер, толкающий бактерию, направление ее поступательного движения будет отличаться от первоначального. Таким способом бактерия может изменять направление своего движения. Электромоторы бактерий являются устройствами, которые в качестве источника энергии используют разность протонных потенциалов на цитоплазматической мембране. Мотор состоит из ротора, статора и некоторых вспомогательных белковых субъединиц, выполняющих роль подшипника, внутри которого вращается стержень ротора.
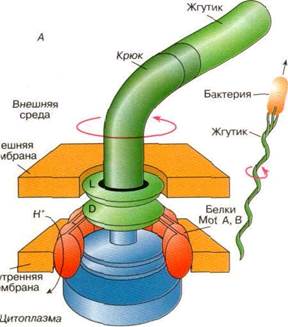
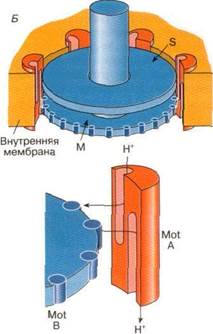
Рис. 1. А - схематическое изображение электромотора, вращающего жгутики бактерий. Центры двух соосных дисхов (М и S) соединены с вращающимся стержнем, выступающим наружу. На периферии диска М находятся моторные белки Mot В. Белки Mot А встроены в мембрану и примыкают к краям дисков М и S;
Б - схема возможного расположения субъединиц Mot А и Mot В, образующих каналы, через которые протоны из периплазматического пространства переносятся в цитоплазму бактериальной клетки Вращающий момент, вызывающий поворот ротора мотора, возникает за счет взаимодействия субъединиц Mot В с белковыми субъединицами Mot A, расположенными на статоре электромотора
Подвижными узлами бактериального электромотора являются два соосных диска (называемые М- и S-дисками), центры которых соединены с вращающимся стержнем, выступающим наружу. На периферии диска М находятся многочисленные копии белка, названного Mot В. Несколько копий белка Mot А, входящего в состав статора, встроены в мембрану и примыкают к краям дисков М и S. Механизм генерации силы, приводящей ротор во вращение, по-видимому, имеет ту же природу, что и в случае АТР-синтазы. Вращающий момент возникает за счет взаимодействия субъединиц Mot В с белковыми субъединицами Mot А, расположенными на статоре электромотора. Считается, что в состав субъединицы Mot А входят два несоосных протонных полуканала. Подобно протонному каналу АТРФ-синтазы, путь переноса протонов через мембрану проходит через протонные полуканалы субъединиц Mot A и Mot В. В результате переноса протонов через белки Mot А и Mot В, направленного внутрь бактериальной клетки, происходит вращение ротора. Один полный оборот ротора связан с переходом через мембрану около 1000 протонов.
Рассмотренными примерами не ограничивается все разнообразие вращающихся моторов, которые встречаются в природе. Существуют бактерии, у которых АТФ-синтазы используют энергию не протонного, а натриевого потенциала.
|
|
|
|
|
Дата добавления: 2015-05-29; Просмотров: 1194; Нарушение авторских прав?; Мы поможем в написании вашей работы!