КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Primary Lymphoid Organs
|
|
|
|
Introduction
Cutaneous-Associated Lymphoid Tissue
Secondary Lymphoid Organs
Lymphatic System
Primary Lymphoid Organs
Introduction
Organs of the Immune System
Lecture 1.3 Structure and function of the immune system
Plan:
A number of morphologically and functionally diverse organsand tissues have various functions in the developmentof immune responses. These can be distinguished by functionas the primary and secondary lymphoid organs (Figure1). The thymus and bone marrow are the primary (orcentral) lymphoid organs, where maturation of lymphocytestakes place. The lymph nodes, spleen, and various mucosalassociatedlymphoid tissues (MALT) such as gut-associatedlymphoid tissue (GALT) are the secondary (or peripheral)lymphoid organs, which trap antigen and provide sites formature lymphocytes to interact with that antigen. In addition, tertiary lymphoid tissues, which normally containfewer lymphoid cells than secondary lymphoid organs, canimport lymphoid cells during an inflammatory response.Most prominent of these are cutaneous-associated lymphoidtissues. Once mature lymphocytes have been generated in theprimary lymphoid organs, they circulate in the blood and lymphatic system, a network of vessels that collect fluid thathas escaped into the tissues from capillaries of the circulatorysystem and ultimately return it to the blood.
Immature lymphocytes generated in hematopoiesis mature and become committed to a particular antigenic specificity within the primary lymphoid organs. Only after a lymphocyte has matured within a primary lymphoid organ is the cell immunocompetent (capable of mounting an immune response). T cells arise in the thymus, and in many mammals – humans and mice for example – B cells originate in bone marrow.
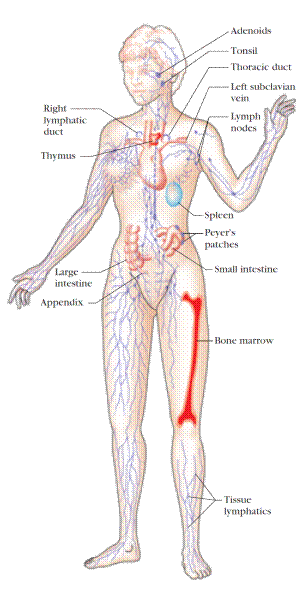
Figure 1. The human lymphoid system. The primary organs (bone marrow and thymus) are shown in red; secondary organs and tissues, in blue. These structurally and functionally diverse lymphoid organs and tissues are interconnected by the blood vessels (not shown) and lymphatic vessels (purple) through which lymphocytes circulate. Only one bone is shown, but all major bones contain marrow and thus are part of the lymphoid system. [ Adapted from H. Lodish et al., 1995, Molecular Cell Biology, 3rd ed., Scientific American Books. ]
THYMUS
The thymus is the site of T-cell development and maturation. It is a flat, bilobed organ situated above the heart. Each lobe is surrounded by a capsule and is divided into lobules, which are separated from each other by strands of connective tissue called trabeculae. Each lobule is organized into two compartments: the outer compartment, or cortex, is densely packed with immature T cells, called thymocytes, whereas the inner compartment, or medulla, is sparsely populated with thymocytes.
Both the cortex and medulla of the thymus are crisscrossed by a three-dimensional stromal-cell network composed of epithelial cells, dendritic cells, and macrophages, which make up the framework of the organ and contribute to the growth and maturation of thymocytes. Many of these stromal cells interact physically with the developing thymocytes (Figure 2). Some thymic epithelial cells in the outer cortex, called nurse cells, have long membrane extensions that surround as many as 50 thymocytes, forming large multicellular complexes. Other cortical epithelial cells have long interconnecting cytoplasmic extensions that form a network and have been shown to interact with numerous thymocytes as they traverse the cortex.
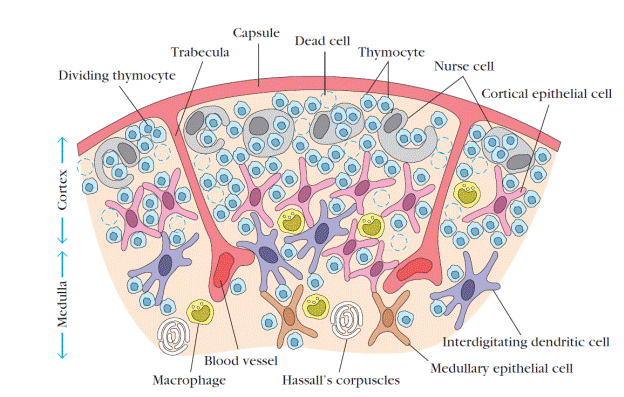
Figure 2. Diagrammatic cross section of a portion of the thymus, showing several lobules separated by connective tissue strands (trabeculae). The densely populated outer cortex is thought to contain many immature thymocytes (blue), which undergo rapid proliferation coupled with an enormous rate of cell death. Also present in the outer cortex are thymic nurse cells (gray), which are specialized epithelial cells with long membrane extensions that surround as many as 50 thymocytes. The medulla is sparsely populated and is thought to contain thymocytes that are more mature. During their stay within the thymus, thymocytes interact with various stromal cells, including cortical epithelial cells (light red), medullary epithelial cells (tan), interdigitating dendritic cells (purple), and macrophages (yellow). These cells produce thymic hormones and express high levels of class I and class II MHC molecules. Hassalls corpuscles, found in the medulla, contain concentric layers of degenerating epithelial cells. [ Adapted, with permission, from W. van Ewijk, 1991, Annu. Rev. Immunol. 9: 591, © 1991 by Annual Reviews. ]
The function of the thymus is to generate and select a repertoire of T cells that will protect the body from infection. As thymocytes develop, an enormous diversity of T-cell receptors is generated by a random process that produces some T cells with receptors capable of recognizing antigen-MHC complexes. However, most of the T-cell receptors produced by this random process are incapable of recognizing antigen-MHC complexes and a small portion react with combinations of self antigen-MHC complexes. The thymus induces the death of those T cells that cannot recognize antigen-MHC complexes and those that react with self-antigen-MHC and pose a danger of causing autoimmune disease. More than 95% of all thymocytes die by apoptosis in the thymus without ever reaching maturity.
THE THYMUS AND IMMUNE FUNCTION
The role of the thymus in immune function can be studied in mice by examining the effects of neonatal thymectomy, a procedure in which the thymus is surgically removed from newborn mice. These thymectomized mice show a dramatic decrease in circulating lymphocytes of the T-cell lineage and an absence of cell-mediated immunity. Other evidence of the importance of the thymus comes from studies of a congenital birth defect in humans (DiGeorge’s syndrome) and in certain mice (nude mice) in which the thymus fails to develop. In both cases, there is an absence of circulating T cells and of cell-mediated immunity and an increase in infectious disease.
Aging is accompanied by a decline in thymic function. This decline may play some role in the decline in immune function during aging in humans and mice. The thymus reaches its maximal size at puberty and then atrophies, with a significant decrease in both cortical and medullary cells and an increase in the total fat content of the organ. Whereas the average weight of the thymus is 70 g in infants, its age-dependent involution leaves an organ with an average weight of only 3 g in the elderly (Figure 3).
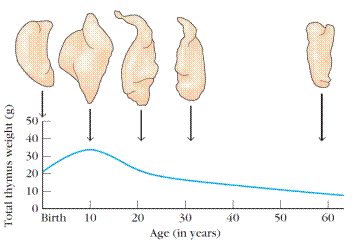
Figure 3. Changes in the thymus with age. The thymus decreases in size and cellularity after puberty.
A number of experiments have been designed to look at the effect of age on the immune function of the thymus. In one experiment, the thymus from a 1-day-old or 33-monthold mouse was grafted into thymectomized adults. (For most laboratory mice, 33 months is very old.) Mice receiving the newborn thymus graft showed a significantly larger improvement in immune function than mice receiving the 33-month-old thymus.
BONE MARROW
In humans and mice, bone marrow is the site of B-cell origin and development. Arising from lymphoid progenitors, immature B cells proliferate and differentiate within the bone marrow, and stromal cells within the bone marrow interact directly with the B cells and secrete various cytokines that are required for development. Like thymic selection during T-cell maturation, a selection process within the bone marrow eliminates B cells with self-reactive antibody receptors. Bone marrow is not the site of B-cell development in all species. In birds, a lymphoid organ called the bursa of Fabricius, a lymphoid tissue associated with the gut, is the primary site of B-cell maturation. In mammals such as primates and rodents, there is no bursa and no single counterpart to it as a primary lymphoid organ. In cattle and sheep, the primary lymphoid tissue hosting the maturation, proliferation, and diversification of B cells early in gestation is the fetal spleen. Later in gestation, this function is assumed by a patch of tissue embedded in the wall of the intestine called the ileal Peyer’s patch,which contains a large number (>1010) B cells. The rabbit, too, uses gut-associated tissues such as the appendix as primary lymphoid tissue for important steps in the proliferation and diversification of B cells.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 2025; Нарушение авторских прав?; Мы поможем в написании вашей работы!