КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
См. Даркшевич,Ueber die hintere Komrnissur, Neurol. Zbl., 1884. 2 страница
|
|
|
|
Заметим здесь, что некоторыми авторами, как Вестфалем и Зимерлингом2, под названием nucl. ventralis post. nucl. oculo-motorii опсывалось содержащее большие многоугольные клетки ядро, расположенное несколько ниже или позади главного ядра и лежащее частью с тылу, частью внутри заднего продольного пучка. Ядро это, как мы впоследствии увидим, в действительности принадлежит боковому нерву и не имеет ничего общего с п. oculomotorius.
Большая часть корешковых волокон п. oculo-motorii возникает из ядер той же стороны и направляется частью в брюшном, частью в наружно-брюшном направлении к месту выхода из мозга; меньшая часть волокон п. oculomo-torii подвергается перекрещиванию между тыльными ядрами и, следовательно, зозникает из ядер противолежащей стороны (рис.7).
К перекрещивающимся волокнам принадлежат собственно задние корешковые волокна п. oculomo-torii, на что в последнее время обратил внимание Бернгеймер; к неперекрещивающимся принадлежат все располагающиеся более кпереди. Первые берут начало в задней половине главного ядра, вторые — в передней его половине, в срединном ядре и в обоих добавочных ядрах.
Задний продольный пучок, несомненно, скрывается частью своих волокон в ядрах п. oculomotorii, преимущественно в глав-
2 Arch. f. Psych., Bd. XXIJ.
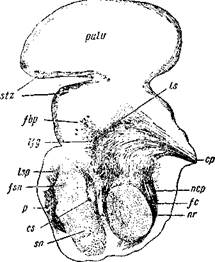
Рис. 6. Срез из мозга несколько недельного младенца на уровне красного ядра pulv — pulvinar thalami; stz — str. zonale.
fbp— волокна задней ручки; //g—волокна петлевого слоя, начинающие рассеиваться при переходе в thalamus; lsp— волокна петлевого слоя, перешедшие в нижний этаж мозговой ножки; fsn— волокна subst. nlgrae; р— пирамидный пучок; «—нижний отдел corp. subtha-lamici; sn— subst.nigra; nr—n. ruber; fc — волокна центрального пучка покрышки; пер— ядро задней спайки; ср — задняя спайка; Is— пучок, выходящий из ядра заднего двухолмия к зрительному бугру. Окраска по Weigert'y.
О В. В. Бехтерев
ndin
ndlll
ном и, может быть, в срединном ядре. Переход волокон заднего продольного пучка в добавочные ядра для меня представляется пока вопросом.
Остальная часть заднего продольного пучка, не скрывающаяся в ядрах п. oculomotorii, на уровне передних отделов главного ядра, начинает постепенно отодвигаться в тыльно-на-ружном направлении по краю центрального серого вещества и вступает в вышеуказанное ядро, принадлежащее брюшному отделу задней спайки. Таким образом, часть заднего продольного пучка является, несомненно, продолжением брюшного отдела задней спайки, другая же часть имеет ближайшее отношение к ядрам п. oculomotorii.
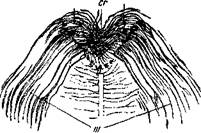
Рис. 7. Препарат из 7-месячного человеческого плода.
///-корешки п. oculomotorii;' nd ///—заднее ядро п. oculomotorii; cr — перекрест корешковых волокон п. oculomotorii между задними ядрами. Окраска по Weigert'y.
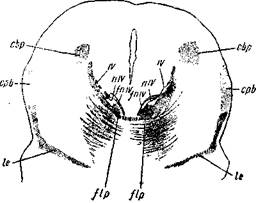
Рис. 8. Срез из мозга 6-месячиого человеческого плода.
cbp — ядро заднего двухолмия; срЬ— corp. parabigeminium или nucl. iemnisci lateralis; lc— наружный отдел главной петли, содержащий волокна из ядер клиновидных пучков; flp — задний продольны.й пучок; IV — корешки п. abducentis; nIV— ядро п. abducentis; fniv — волокна, выходящие из ядра п. abducentis в область заднего продольного пучка.
С другой стороны, к ядрам п. oculomotorii подходят волокна глубокого отдела переднего двухолмия, веерообразно расходящиеся в области серого вещества последнего. Большая часть этих волокон образует с брюшной стороны от центрального серого
вещества так называемый фонтановидный перекрест и спускается затем в нисходящем направлении во внутренних отделах сете-видного образования к спинному мозгу. Меньшая же часть, несомненно, вступает в ядра п. oculomotorii. Таким образом, часть волокон заднего продольного пучка, возникающего из ядер п. oculomotorii, является, по крайней мере частью, продолжением волокон глубокого отдела переднего двухолмия.
Что касается ядер блокового нерва, то в этом отношении до позднейшего времени существовали не незначительные разноречия в литературе; так, например, за главное ядро п. trochlearis неко торыми авторами и, между прочим, Вестфалем и Зимер-лингом признавалось ядро, содержащее довольно большие закругленные клеточные элементы и располагающееся на уровне заднего двухолмия внутри центрального серого канала в тыльном и отчасти в тыльно-наружном направлении от заднего продольного пучка. С другой стороны, кругловатая группа мелких клеточных элементов, прилежащая к хвостовой части выше описанного ядра, была описана Вестфалем под названием заднего мел ко клеточного ядра п. trochlearis. В последнее время против принадлежности того и другого ядра к trochlearis высказался Кауш \ который за истинное ядро п. trochlearis признает ядро расположенное на тех же уровнях внутри заднего продольного пучка и признаваемое некоторыми авторами за п. centralis posterior п. oculomotorii (см. выше). Эти исследования Кауша, однако, не остались без возражения, в силу чего, мне кажется в этом отношении могут иметь значение указания, сделанные на основании эмбриологического метода.
Вопрос о принадлежности вышеупомянутого ядра к п. trochlearis может быть с точностью выяснен на зародышевых мозгах в силу того обстоятельства, что корешковые волокна, как известно,
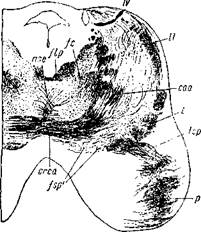
Рис. 9. Косвенный разрез через мозговой ствол сверху и сзади вперед и вниз из мозга несколько недельного младенца.
nse— область верхняя внутреннего центрального ядра; flp— задний продольный пучок; !с— центральный пучок покрышки; IV— корешки блокового нерва после перекреста над aq. Sylvii; II— боковая петля; /—главная петля; lsp — волокна петлевого слоя, переходящие в мозговую ножку; р— пира мидиый пучок; /sp'— рассеянные пучки тонких волокон внутри петлевого слоя; сгса — перекрест волокон передней ножки мозжечка. Окраска по Weigert'y.
К a u s с h, Neurol. Zbl., 1894, № 14.
6'.'
развиваются очень рано, волокна же окружающие центральное серое вещество, исключая задний продольный пучок, развиваются в более позднем возрасте. Пользуясь подходящим материалом из человеческих зародышей, я имел полную возможность убедиться в принадлежности вышеуказанного ядра, заложенного внутри заднего продольного пучка к корешкам п. troch-learis, которые могут быть непосредственно прослежены в этом ядре (рис. 8, nIV).
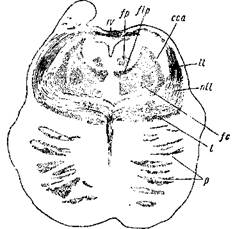
Рис. 10. Срез позади четверохолмия из мозга несколько недельного младенца.
IV — корешки блокового нерва, перекрещивающиеся над aq. Sylvii; fp — область заднего пучка Schutz'a; flp — задний продольный пучок; сса — передняя ножка мозжечка; II— боковая петля; nil— ядро боковой петли; fc— центральный пучок покрышки; /—главная петля; р— пирамидный пучок.
Ввиду того, что некоторые авторы высказывали сомнение в полном перекресте волокон блокового нерва позади заднего двухолмия, заметим здесь, что метод развития в выяснении этого вопроса оказывает незаменимые услуги и ставит вне сомнения существование полного перекреста волокон блокового нерва.
Ход этого нерва на всем протяжении внутри мозга крайне легко проследить на зародышевых мозгах, где его волокна представляются мякотными еще в том возрасте, когда соседние части мозга или вовсе не содержат мякотных волокон, или содержат их лишь в малом количестве. На срезах из таких мозгов можно убедиться, что волокна блокового нерва образуют полный пере-кргст внутри переднего паруса, не перекрещенных же волокон не существует вовсе (рис. 9 и 10, IV).
После перекреста волокна блокового нерва проходят на некотором протяжении в виде круглых пучков снутри от нисходя-
щего корешка тройничного нерва, после чего, отклоняясь дугообразно внутрь и в брюшном направлении, они достигают ядра своей стороны, о котором мы упомянули выше (рис. 8, nIV). Что касается ядер п. abducentis, располагающихся в тыльных частях моста под местом загиба корешка личного нерва (рис. И, riVI), то на зародышевых мозгах можно убедиться, что они посылают от себя волокна лишь в корешки соответствующей стороны, следовательно, не существует предполагаемого некоторыми ав-
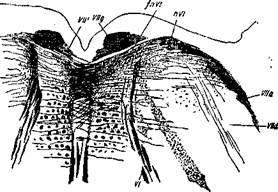
Рис. 11. Разрез на уровне ядра п. abducentis из человеческого плода 33 см длины.
'Vila— восходящая часть корешка facialis; Vlld — нисходящая часть корешка facialis; nVI — ядро п. abducentis; fnVI— волокна, выходящие из ядра п. abducentis к заднему продольному пучку противолежащей стороны; V//g—колено корешка п. facialis; VII' —волокна п. facialis, направляющиеся к противоположной стороне; VI — корешки п. abducentis. Окраска no Weigert'y-
торами неполного перекрещивания корешковых волокон отводящего нерва, что вполне согласно с новейшими исследованиями Бергмана, произведенными по методу перерождения1.
Относительно взаимной связи ядер п. abducentis и п. oculomotorii, как известно, до сих пор остается еще много невыясненного. Правда, уже Дюваль и Лаборд указали на существование волокон, проходящих под дном IV желудочка от ядра п. abducentis одной стороны к ядру п. oculomotorii другой стороны. Однако эти указания не могли быть подтверждены при позднейших исследованиях, а потому в виду доказанного перекрещивания части корешковых волокон п. oculomotorii некоторые авторы принимали, что между тем и другим ядром существует прямая связь при посредстве волокон, проходящих в заднем продольном пучке.
1 Bergmann, Jahrb. f. Psych., Bd. XI.
На основании своих исследований зародышевых мозгов и мозгов новорожденных я убедился, однако, что из каждого ядра отводящего нерва, действительно, отходят в довольно значительном числе волокна внутрь к заднему продольному пучку, который они проникают насквозь и затем, перекрестившись во шве, достигают заднего продольного пучка другой стороны (рис. 11, fnVI). Более чем вероятно, что эти волокна именно и служат для связи ядра отводящего нерва одной стороны с ядром п. oculomotorii другой стороны.
Равным образом на основании эмбриологического материала нетрудно убедиться, что и от ядра п. trochlearis направляется небольшой пучок волокон в область заднего продольного пучка своей стороны (рис. 8, fn.IV); пучок этот, подобно предыдущим волокнам, очевидно, также служит для сочетательной деятельности глазных яблок.
Наконец, и между ядрами п. oculomotorii обеих сторон имеется сочетательная система волокон, которая, без сомнения, также имеет известное значение при совместной деятельности глазных яблок.
05 ОСОБОМ ВНУТРЕННЕМ ПУЧКЕ БОКОВЫХ СТОЛБОВ
Как известно, часть белого вещества боковых столбов, расположенная на границе с серым веществом, была выделена Флек-сигом под названием пограничного слоя. Последний, по Флек-сигу, в верхней шейной области располагается кнаружи от переднего рога, простираясь от передней его группы до заднего конца рога. В шейном утолщении он занимает угол между передним и задним рогом. В грудной области его передняя часть проходит снова вперед и образует пучок, располагающийся снаружи переднего рога; но он представляется здесь тоньше, нежели в верхней шейной области. В поясничном же утолщении отношения этого пучка еще недостаточно выяснены. Что касается состава волокон пограничного слоя, то в нем предполагались частью волокна короткого протяжения, частью волокна из кларковых столбов, частью, наконец, волокна из задних корешков. Не подлежит, впрочем, сомнению, что волокна задних корешков не входят в состав пограничного слоя, так как перерезка задних корешков не влечет за собой перерождения волокон пограничного слоя.С другой стороны, и волокна из столбов Кларка лишь пересекают область пограничного слоя на своем пути к мозжечковому пучку, но не входят в состав продольных волокон.
Уже много лет тому назад, еще во время заведывания мною психофизиологической лабораторией Казанского университета, на основании исследований по методу развития мне удалось выделить в вышеуказанном пограничном слое один пучок, располагающийся в более задних отделах этого слоя, который я назвал внутренним пучком боковых столбов. Об этом пучке я говорю хотя и сжато, но вполне определенно уже в первом издании своих «Проводящих путей», вышедшем в 1893 г. на русском языке и в 1894 г. в переводе на немецкий язык. Равным образом об этом пучке я говорю теми же словами и во втором издании «Проводящих путей», вышедшем на русском языке в 1896 г. 1.
1 См. Проводящие пути спинного и головного мозга. Руководство к изучению внутр~"иих связей мозга, изд. Риккера, 1896.
Из этого издания я заимствую здесь следующие строки (см. стр. 99 «Проводящих путей мозга»).
«Основной пучок кзади распространяется частью также и на область, известную под названием пограничного слоя. В последнем, однако, следует различать особый пучок, который в силу его положения я называю внутренним пучком. Волокна этого пучка развиваются позднее волокон основного пучка, но ранее волокон других соседних пучков, благодаря чему он легко может быть выделен на зародышевых мозгах соответствующего возраста (рис. 48 и 49)»1.
«Рассматриваемый пучок с ясностью обнаруживается уже в грудной области спинного мозга, где он располагается непо средственно снаружи от серого вещества между боковым и задним рогом, занимая собою, таким образом, часть пространства, заключающегося между пирамидным пучком и серым веществом спинного мозга, далее же кверху пучок этот продвигается более кпереди, располагаясь все время вблизи серого вещества».
Что касается происхождения волокон внутреннего пучка, то я высказался в том же издании следующим образом:
«Надо думать, что он имеет отношение к клеточным элементам, располагающимся в более задних областях серого вещества, и состоит из коротких волокон, так как перерождение его у человека до сих пор еще не наблюдалось. Впрочем, при перерезке спинного мозга у кролика мне случалось наблюдать восходящее перерождение этого пучка на коротком протяжении кверху от места разрушения».
Ввиду всего вышеуказанного нельзя не признать совершенно ошибочным утверждение А. Брюса2, будто бы до сих пор не было сделано подразделения пограничного слоя на составные части. Последний автор, между прочим, опубликовал случай амиотро-фического бокового склероза, в котором вместе с перерождением боковых пирамидных пучков и клеток передних рогов обнаруживалось также перерождение волокон основного пучка передне-боковых столбов спинного мозга и одновременно с тем перерождение волокон передней части пограничного слоя; между тем как волокна его задней части, стоящей в связи с клетками задних рогов, представлялись неперерожденными
Благодаря этому автор описывает заднюю часть пограничного слоя в виде особого пучка, широкий конец которого располагается в углу, образованном передним и задним рогом, задний же конец, постепенно истончаясь, протягивается почти до верхушки заднего рога.
1 Упомянутые здесь рнсункн приведены там же, на стр. 99 «Проводящих путей мозга».
2 А. В г и с е, Revue neurol., 1896, № 23.
Сравнивая это описание и рисунки автора, я должен признать, что они относятся к той именно области пограничного слоя, которую я описал уже несколько лет назад под названием «внутреннего пучка боковых столбов».
Таким образом, в исследовании А. Брюса, основывающимся на методе перерождения, я нахожу полное подтверждение впервые выделенного мною по методу развития в задней части пограничного слоя особого внутреннего пучка боковых столбов спинного мозга.
ОБ ОСОБОМ ЯДРЕ СЕТЕВИДНОГО ОБРАЗОВАНИЯ НА УРОВНЕ ВЕРХНИХ ОТДЕЛОВ ВАРОЛИЕВА МОСТА
В последнее время, благодаря исследованиям Роллера, моим, Миславского и др., в сетевидном образовании, кроме уже ранее хорошо известных образований (как например, верхние оливы и пр.), удалось выделить несколько новых или мало известных ранее гнезд серого вещества, в числе которых, придерживаясь восходящего топографического расположения, мы назовем: 1) описанное Миславским ядро переднего столба, или дыхательное ядро, заложенное в продолговатом мозгу на уровне корешков п. hypoglossi частью кнутри, частью кнаружи от последних;
2) нижнее центральное ядро Роллера, расположенное в продолговатом мозгу над головным или верхним отделом нижних олив;
3) описанное мною на уровне моста сетчатое ядро покрышки, расположенной над внутренней частью петлевого слоя; 4) описанное мною верхнее центральное ядро, лежащее по обе стороны шва на уровне заднего двухолмия; 5) описанное мною же безымянное ядро, заложенное на уровне переднего двухолмия снутри и стылу от петлевого слоя, представляющегося на этом уровне на поперечных срезах в виде серповидного пучка, и, наконец, 6) расположенное еще выше небольшое конической формы ядро поперечного пучка мозговой ножки.
Кроме этих гнезд серого вещества, в сетевидном образовании может быть выделено еще одно важное гнездо на уровне моста тотчас позади заднего двухолмия, заложенное в глубоких частях покрышки г.
Ядро это мне удалось выделить прежде всего на последовательном ряде окрашенных пикрокармином срезов из мозгового ствола кошки; у которой оно состоит из редко расположенных многоотростчатых больших нервных клеток, различаемых на
1 Рисунок этого ядра и топографическое отношение его к другим соседним ядрам можно найти в первом томе моих «Проводящих путей мозга», СПБ, 1896, стр. 148, рис. 91, под буквами пс se; об этом ядре я упоминаю в том же издании на стр. 143.
окрашенных срезах даже невооруженным глазом в виде мелких точек. Таким образом, по своей величине клетки описываемого ядра ничуть не уступают двигательным клеткам передних рогов спинного мозга, а некоторые из его клеточных элементов даже превосходят последние.
В гистологическом отношении описываемое ядро, состоящее из больших многоотросчатых клеток, заложенных среди белого вещества, довольно близко напоминает собой нижнее центральное ядро и подобно последнему располагается в тех частях сете-видного образования, которые являются продолжением основных пучков боковых столбов. В силу этого ядро это может быть названо наружным верхним центральным ядром, или просто верхним центральным ядром, тогда как описанное мною ранее верхнее центральное ядро в отличие от предыдущего может быть названо внутренним верхним центральным ядром или еще лучше срединным ядром.
Вышеуказанное ядро на соответствующем уровне найдено мною, между прочим, и в мозгу собаки, а также и у человека; но у последнего клетки этого ядра не так значительны по своей величине, вследствие чего и самое ядро обозначается далеко не столь резко, как в мозгах кошки и собаки.
Что касается до связей этого ядра, то пока можно сказать лишь, что подобно нижнему центральному ядру оно располагается на пути волокон, являющихся продолжением основного пучка боковых столбов спинного мозга и по всей вероятности стоит в ближайшем соотношении с волокнами последних.
О РАЗВИТИИ КЛЕТОК МОЗГОВОЙ КОРЫ У ЧЕЛОВЕКА х
(Краткое сообщение)
При исследовании зародышевых мозгов человека по методу Гольджи в заведываемой мною лаборатории удалось хорошо окрасить нервные элементы мозговой коры как в период первоначального образования клеток последней, так и в период дальнейшего их роста и развития. На основании этих препаратов я могу сделать следующие выводы относительно развития нервных клеток мозговой коры.
1. Первоначальному образованию клеток мозговой коры на место зародышевых элементов предшествует рост окрашивающихся по Гольджи в черный цвет особых нитей, resp. волокон, выходящих густыми рядами со стороны также окрашенного в черный цвет поверхностного слоя, углубляющихся в зародышевую мозговую ткань и проникающих между зародышевыми тельцами. В наиболее раннем периоде встречаются лишь небольшой длины первичные нити, проникающие в наиболее наружные отделы мозговой коры, позднее же эти нити представляются уже значительно большей длины и достигают более глубоких ее слоев.
2. Самые клетки обязаны своим развитием тому, что вышеуказанные первичные нити при своем развитии, проникая между зародышевыми тельцами, образуют на своем пути изгибы в виде колена или складки вокруг зародышевых телец. Последние при постепенном увеличении колена или изгиба нитей обволакиваются ими по всей своей окружности, образуя, таким образом, закругленный, чаще всего овальный остов будущей нервной клетки, обращенная же кнаружи часть первичной нити становится верхушечным отростком клетки, являющимся, таким образом, по своему развитию наиболее ранним из всех вообще клеточных отростков.
3. Зародышевое тельце, неокрашивающееся по методу Гольджи, служит при выше описанном развитии нервной клетки
1 Доклад, сделанный в научном собрании врачей клиники душевных и нервных болезней в С.-Петербурге 15/IV 1896 г. с демонстрацией препа* ратов.
к образованию ее ядра, вещество же нити, обволакивающее зародышевое тельце, служит к образованию клеточной протоплазмы и ее отростков. Благодаря этому во всех клетках наиболее раннего возраста клеточные ядра представляются совершенно неокрашенными, количество же окружающей его протоплазмы по сравнению с размерами ядра оказывается весьма незначительным.
4. В дальнейшем периоде развития от овального остова тела клетки при их росте выступают в виде толстых шипов боковые отпрыски, развивающиеся в протоплазменные отростки, тела же клеток, благодаря этому, становятся угловатыми или близкими по своей внешней форме к пирамидкам. Вместе с тем и узловатости первичных нитей при дальнейшем росте служат началом шипо-образных выступов, которые со временем становятся боковыми побегами верхушечных протоплазменных отростков. Коллатера-ли осевых цилиндров вырастают всегда позднее самих цилиндров на месте четкообразных вздутий последних.
5. Богатое развитие протоплазменных клеточных отростков и их дальнейших разветвлений есть также продукт позднейшего развития клеток, так как более молодые клетки, имея уже сравнительно хорошо развитый верхушечный отросток, почти вовсе лишены как основных, так и боковых протоплазменных отростков или же имеют лишь весьма ограниченное количество основных коротких отростков (согласно Маджини г, Кахалю2 и др.3). Лишь со временем, вместе с ростом тела клетки, развиваются или удлиняются основные отростки и затем ветвится верхушечный отросток, как это мы видим у вполне развитых клеток.
6. Что касается так называемых грушевидных придатков (appendices pyriformes), то они совершенно отсутствуют у более молодых клеток и развиваются на клеточных отростках лишь в позднейшее время, когда клетки уже приобретают вид взрослых. Ранее всего и притом в большом изобилии они развиваются, по-видимому, на верхушечном отростке как более раннем по своему развитию. С развитием протоплазменных отростков число этих придатков все более и более увеличивается4.
7. Различные области мозговой коры развиваются далеко не в одинаковое время. Тогда как в одних областях, например, в чувство-двигательной, мы встречаем уже ясные зачатки клеток, в других областях (например, в subiculeum аммонова рога) мы
1 М a g i π i, Sur la neurologic et les cellules nerveuses cerebrales chez le foetus, Arch. ital. de biol., IX, p. 59.
2 Ramon у Cachal, Sur la structure de 1' ecorce cerebrale de quel-ques mamumiferes, La cellule, 1891, v. VII.
3 К аналогичным результатам пришла в позднейшее время и Стефанов-ская на основании исследований, произведенных над молодыми крысами (М. S t e f a η owsk a, Evolution des cellules nerveusses corticales, Bru-xelles, 18ОД).
4 M. Стефановская, там же, стр. 33 н следующие.
находим лишь первичные нити без всяких следов образования нервных клеток. Равным образом и клетки различных слоев одной и той же области мозговой коры развиваются неодновременно: клетки более глубоких слоев, например, слоя больших пирамид, развиваются ранее клеток более поверхностных слоев.
8. В общем клетки мозговой коры принадлежат по своему развитию к наиболее поздним клеточным элементам центральной нервной системы, так как они представляются еще в зачаточном состоянии в то время, как клетки мозговых узлов представляются уже сравнительно хорошо сформировавшимися, а клетки спинного мозга уже»вполне развитыми.
II
ТРУДЫ ПО ФИЗИОЛОГИИ
ОБ ОТПРАВЛЕНИИ ЗРИТЕЛЬНЫХ БУГРОВ
В физиологии нервной системы нет более темной области, как отправление зрительных бугров. Различные авторы высказали в этом отношении самые разноречивые мнения, что, повиди-мому, давало возможность предполагать о весьма многосторонней роли упомянутых узлов в мозговой деятельности. Между тем до настоящего времени только отношение этих узлов к зрению может считаться более или менее прочно установленным, что, впрочем, очевидно уже из анатомических данных. Все же остальные указания, сделанные теми или другими авторами, относительно значения зрительных бугров в отправлениях нервной системы оспаривались с самых различных сторон и далеко не могли послужить к действительному выяснению отправления этой важной части мозговых полушарий.
Мы считаем здесь необходимым вкратце упомянуть только о наиболее существенных данных, встречающихся в физиологической литературе зрительных бугров.
Начиная с Роландо, многие авторы (из более новых Ренци1, Нотнагель и Ферье2) упоминают о слепоте противуположного глаза, наступающей у животного вслед за повреждением зрительного бугра, даже и в том случае, если разрушению подверглась только передняя его часть. Вместе с тем, некоторые из физиологов думали также установить влияние зрительных бугров и на движение зрачка. Так, Лонже3 при разрушении бугра наблюдал неподвижность зрачка при прямом раздражении светом. Между тем Ренци указывает, что подобное явление наступает только при разрушении передней части узла, тогда как возможно полное разрушение задней его части вызывает продолжительное и сильное расширение противуположного зрачка. Последнее наблюдал также и Ферье в опыте над обезьяной.
Далее, приводили множество данных, говорящих в пользу влияния зрительных бугров на различные роды чувствительности.
1 Renzi, Saggio di fisiologia, Annalt univers. di medicina, m. 186.
2 Ferrier, Die Funktionen des Gehirns; нем. перев. Obersteiner'a, Braunschweig, 1879, стр. 265 и ел.
8 Longet, Anatomie et physiologie du systeme' nerveux, т. 1, стр. 500 и ел. У
6 В. М. Бехтерев 81
Утверждали, например, что, помимо значения бугра для зрения, он влияет также на слух, обоняние, вкуси кожные ощущения.
Представителями взгляда, допускающего чувствительную функцию зрительных бугров, могут считаться Ренци, Фурнье1, Ферье и в особенности Люи2. Мнение этих авторов основывается, главным образом, на том, что при экспериментальных 'исследованиях, в особенности над низшими животными, вслед за разрушением бугра нередко наблюдали притупление или полное прекращение чувствительности на всей противуположной половине тела. Люи в пользу своего взгляда приводит также несколько патологических наблюдений.
В литературе существуют также весьма многосторонние указания о влиянии зрительных бугров на двигательную сферу. Оставляя в стороне взгляды старых авторов (Сосерот, Серре и др.), локализировавших в этих узлах центры для движения передних конечностей, мы находим много указаний на появление вслед за разрушением бугров различных вынужденных движений. Так, Мажанди3 при своих опытах наблюдал круговые движения на сторону поврежденного узла, а Лонже — в противуположной направлении; между тем Шифф4 утверждает, что только повреждение задней части бугра, подобно повреждению мозговой ножки, вызывает кружение животного на здоровую сторону (согласно с мнением Лонже); повреждение же передней трети узла сопровождается кружением животного в противупо-ложную сторону (согласно синением Мажанди). Впрочем указания Ренци более согласуются с мнением Лонже. Кроме того, Ренци в опытах над птицами с разрушением мозговых полушарий вместе с зрительными буграми наблюдал полную неспособность животных стоять, ходить и летать, несмотря на то, что они могли еще производить отдельные движения и махали крыльями при бросании их на воздух.
Наконец, не очень давно были опубликованы опыты Нотнаге-ля (над кроликами), в которых разрушение бугров производилось впрыскиванием хромовой кислоты. Существенные результаты этих опытов состояли в том, что оперированные животные не представляли никаких явлений двигательного или чувствительного паралича, но. отличались от здоровых тем, что при пассивном вытягивании или передвижении их передних конечностей-оки долгое время сохраняли новые положения своих членов. Принимая, что исправление положения членов у нормальных животных выполняется отраженным путем под контролем мышечного чувства, ав-
|
|
|
|
|
Дата добавления: 2015-06-26; Просмотров: 284; Нарушение авторских прав?; Мы поможем в написании вашей работы!