КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структурно-функциональные нарушения ИК
|
|
|
|
Функциональные состояния ИК
Функции ИК
Главная функция ИК - обеспечивать управляемое перемещение ионов через мембрану.
В зависимости от проходящих через них ионов ИК подразделяют на натриевые, калиевые, кальциевые, хлорные, протонные (водородные).
Функции ИК:
1. Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление и защелачивание.
3. Регуляция ионного обмена (обмен солей): изменение внутриклеточного ионного состава и концентрации.
4. Создание и изменение мембранных потенциалов: потенциал покоя; в возбудимых клетках - локальные потенциалы, потенциал действия.
5. Проведение возбуждения в возбудимых клетках: обеспечение движения нервных импульсов.
6. Трансдукция в сенсорных рецепторах: преобразование раздражения (стимула) в возбуждение.
7. Управление активностью клетки: за счёт обеспечения потоков вторичного мессенджера - Са2+.
1. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал закрыт и ионы не проходят через него.
3. Активированное. Канал может выполнять свои функции, т.е. открываться и закрываться под действием его регуляторов (управляющих веществ или электрических потенциалов).
4. Инактивированное. Канал не может выполнять свои функции, т.е. открываться и закрываться, он "фиксируется" в каком-то одном состоянии.
5. Блокированное. Канал перекрыт, инактивирован веществом-антагонистом (блокатором), занявшем место управляющего вещества.
6. Модулированное (фосфорилированное). Канал изменяет свои обычные свойства под действием фосфорилирования - присоединения к какому-то его участку фосфатного остатка.
Каналопатии - это группа врождённых заболеваний, вызванных нарушениями в работе ИК. Каждая каналопатия обусловлена патологией соответствующих ИК. В основе патологии лежит либо мутация генов, кодирующих строение белков-каналоформеров, либо производство аутоантител, агрессивных по отношению к собственным ИК.
Функциональная классификация ионных каналов (ИК)
( © Сазонов В.Ф., 2011. © http://kineziolog.bodhy.ru, 2011.)
Как уже говорилось выше, ИК можно классифицировать различным образом:
1. По селективности (степени избирательной проницаемости к определённым ионам). В этом случае мы будем говорить о натриевых, калиевых, хлорных каналах и т.п.
2. По строению (родству их химического строения и происхождения образующих их белков). По строению (структуре) и по происхождению от однотипных генов различные ИК объединяются в отдельные семейства. Например, выделяют три семейства лиганд-активируемых ИК: 1) семейство с пуриновыми рецепторами (АТФ-активируемые), 2) с никотиновыми АХ-рецепторами, ГАМК-, глицин- и серотонин-рецепторами, 3) с глутаматными рецепторами. При этом в одно и то же семейство попадают ИК с разной ионной селективностью, а также ИК с разными управляющими лигандами. Но зато образующие эти каналы белки имеют большое сходство в строении и происхождении.
3. По способу управления их состоянием. В этом случае мы будем говорить о потенциал-управляемых каналах, хемо-управляемых и т.д.
4. По связывающимся с ними лигандам (в том числе веществам-маркёрам) и т.д.
Создание удобной классификации является пока ещё не решённой проблемой. Как указывают Н.Н. Мушкамбаров и С.Л. Кузнецов, (2003), "в отличие от липидов, мембранные белки трудно классифицировать по их структуре. Более перспективно попытаться подразделить эти белки по их функциональной роли. Но и здесь нет законченной системы, т.к. любые попытки её создания наталкиваются на типичные трудности, когда один и тот же белок может быть отнесён к разным группам". Тем не менее, на наш взгляд, функциональная классификация ИК просто необходима для обучения студентов: биологов, медиков, психологов.
В основу предложенной нами функциональной классификации ИК (Сазонов В.Ф., 2011) положен способ управления их деятельностью, а не их селективная проницаемость к определённым ионам или химическое родство образующих их белков-каналоформеров. С этой точки зрения ИК делятся на неуправляемые и управляемые, т.е. либо постоянно открытые, либо открывающиеся-закрывающиеся при определённих воздействиях. Заметим, что большинство ИК являются управляемыми, но различаются между собой по механизмам управления. Воздействие регуляторного (управляющего) фактора на управляемый ИК вызывает конформационные изменения каналообразующих белков, канал открывается и ионы проходят по градиенту концентрации. При этом сам транспорт ионов через такие каналы не приводит к конформационным изменениям канальных белков и зависит только от разности концентраций веществ по обе стороны мембраны.
В одну и ту же функциональную группу нашей классификации могут попасть каналы различного молекулярного строения и с различной селективностью, т.е. пропускающие различные ионы. С другой стороны, сходные по строению и происхождению каналы могут оказаться в разных функциональных группах. Так, например, хлор-селективные ИК могут управляться как лигандами (глицином, ГАМК) и состоять в группе лиганд-управляемых каналов, так и потенциалом мембрнаны и состоять в группе потенциал-управляемых каналов (потенциал-активируемые хлорные каналы ClC).
Студентам
Для понимания электрических процессов, идущих в нервных клетках, формирования электрических потенциалов и нервных импульсов вполне достаточным будет разобраться в первых четырёх видах ионных каналов: 1) неуправляемые постоянно пропускают через себя ионы калия, 2) потенциал-управляемые открываются при деполяризации и начинают в этих условиях пропускать через себя в клетку ионы натрия (в постсинаптических окончаниях и нервных отростках) или же ионы кальция (в пресинаптических окончаниях или рецепторных клетках), 3) хемо-управляемые открываются под действием медиатора и начинают пропускать через себя в клетку ионы натрия, что вызывает деполяризацию в виде возбуждающего постсинаптического потенциала (ВПСП), 4) стимул-управляемые находятся в сенсорных рецепторах (рецепторных клетках или рецепторных нервных окончаниях) и открываются под действием стимула (раздражителя), начиная пропускать через себя ионы натрия, что вызывает деполяризацию в виде рецепторного потенциала.
Виды ионных каналов согласно функциональной классификации:
1. Неуправляемые (независимые). Конечно, это название условно и отражает лишь основное функциональное состояние подобных каналов. Пожалуй, полностью независимых и неуправляемых ИК в мембране просто не существует, и все они так или иначе регулируются. Неуправляемые ИК обычно находятся в постоянно открытом состоянии и обеспечивают постоянный ионный ток через открытую пору канала как в клетку, так и из клетки. Процесс перемещения ионов через такие ИК идёт пассивно за счёт диффузии под действием химических сил (по градиенту их концентрации) и/или электрических сил (по электрическому градиенту зарядов между внутренней и наружной сторонами мембраны).
Если неуправляемые каналы различают вещества только по размеру и пропускают через себя по градиенту концентрации все молекулы меньше определённой величины, т.е. служат фильтрами молекулярных размеров, то их называют "неселективные каналы", или "поры". Селективные каналы, работают избирательно и обеспечивают перенос только определённых ионов. Ионная селективность (избирательность) каналов определяется их диаметром и строением внутренней поверхности канала. Например, катионселективные каналы пропускают только катионы, так как содержат много отрицательно заряженных аминокислотных остатков.
Примеры: калиевые каналы утечки (они участвуют в формировании нервными клетками мембранного потенциала покоя), эпителиальные натриевые каналы ENaCs (они обеспечивают обратное всасывание ионов натрия в почках, прямой кишке, лёгких, потовых железах и пр., также обеспечивают восприятии солёного вкуса вкусовыми рецепторами во рту), анионные каналы эритроцитов (они пропускают анионы Сl-, OH-, HCO3-).
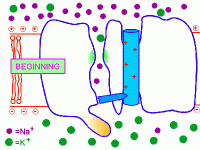
2. Потенциал-управляемые (потенциал-чувствительные, потенциал-зависимые, потенциал-активируемые, voltage-gated). Так, потенциал-управляемые натриевые каналы открываются под действием сдвига электрического потенциала мембраны, превышающего критический уровень деполяризации. Поэтому при достижении определённого порогового уровня деполяризации мембраны они открываются, а при обратном снижении уровня деполяризации - оказываются закрытыми. Важно, что как при химической, так и при фармакологической модификации таких ИК у них сохраняется основной механизм активации и инактивации в ответ на сдвиг мембранного потенциала, что и определяет быстрые изменения катионной проницаемости возбудимых мембран за счёт потенциал-управляемых ИК. Именно такого типа потенциал-управляемые натриевые ИК обеспечивают перемещение нервного импульса по мембране нейрона (смотри: потенциал действия и нервный импульс).
Примеры: т етродотоксин-чувствительные натриевые каналы, потенциал-активируемые К-каналы, калиевые Kdr-каналы задержанного выпрямления, кальциевые каналы пресинаптических окончаний аксонов.
На рисунке справа - условная схема работы потенциал-управляемого ИК (кликните на рисунок, чтобы увидеть процесс в динамике).
3. Хемо -управляемые (хемочувствительные, хемозависимые, лиганд-управляемые, лиганд-зависимые, рецептор-активируемые). Они открываются при связывании с рецепторным участком канала специфического лиганда (управляющего вещества: трансмиттера или его миметика). Такие каналы обычно локализованы в химических синапсах на их постсинаптических мембранах и преобразуют химический сигнал, возникающий за счёт пресинаптического высвобождения нейромедиатора, в постсинаптический электрический локальный потенциал. Смотри: локальный потенциал, 3_3 Синапсы, синапсы, медиаторы и модуляторы.
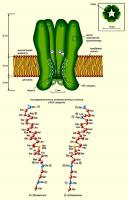 Примеры: каналы с никотиновыми ацетилхолиновыми рецепторами nAChR), серотониновыми рецепторами (5-HT3), глициновыми, ГАМК-рецепторами (GABAA и GABAC).
Примеры: каналы с никотиновыми ацетилхолиновыми рецепторами nAChR), серотониновыми рецепторами (5-HT3), глициновыми, ГАМК-рецепторами (GABAA и GABAC).
На рисунке справа - лиганд-управляемый ионный канал с никотиновым ацетилхолиновым рецептором (никотин - миметик, ацетилхолин - трансмиттер). Канал состоит из 5 субъединиц и поры в центре. Вверху — поперечный разрез этих субъединиц: a1, a2, b, g, d. Внизу — участки субъединиц, образующих «воротную систему» канала. Представлены аминокислотные последовательности М2 a-спирали в b- и d-. Из 5 субъединиц, образующих пору, изображены лишь 4, а ближайшая к нам удалена, чтобы были видны участки М2, облицовывающие ионный канал, и ворота. Видно, что большая часть молекулы белка выходит за пределы внешней поверхности плазматической мембраны, образуя молекулярные рецепторы к лиганду. Каждая из двух a-субъединица содержит связывающий центр для ацетилхолина, следовательно, с рецептором может связаться 2 молекулы лиганда (нейротрансмиттера или нейромиметика). Ворота, находящиеся в пределах поры, открываются при связывании ацетилхолина с рецепторным участком канала. Отрицательно заряженные остатки глутаминовой и аспарагиновой аминокислот (они выделены синим цветом) имеются в обоих концах М2-спиралей, т.е. с двух сторон поры, благодаря чему предотвращается вход в канал анионов, а катионы Na+ и К+ при его закрытом состоянии могут быть связаны уже в самом канале.
4. Стимул-управляемые (механочувствительные, механосенситивные, стретч-активируемые, stretch-activated, протон-активируемые, температурно-чувствительные).
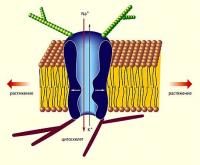
Они открываются под воздействием специфичного и адекватного для них стимула (раздражителя). Такие каналы обеспечивают сенсорное восприятие и располагаются в мембране сенсорных рецепторов.
Пример: механочувствительные ИК рецепторных волосковых клеток, обеспечивающих слуховое восприятие; температурно-чувствительные ИК терморецепторов кожи, обеспечивающие восприятие тепла и холода.
В настоящее время стимул-управляемые механочувствительные ИК обнаружены не только в специализированных механорецепторных структурах, но также и в мембранах бактерий, грибов, растений, позвоночных и беспозвоночных животных. Механочувствительные каналы не только обеспечивают сенсорное восприятие механического раздражения, но также вовлечены в контроль клеточного цикла, регуляцию объёма и роста клеток, секрецию и эндоцитоз.
TRP-каналы в мембране терморецепторов кожи обеспечивают термотрансдукцию, открываясь при различных значениях темпераруры. Они пропускают катионы, особенно ионы кальция.
5. Совместно-управляемые (NMDA-рецепторно-канальный комплекс). Они открываются одновременно как лигандами, так и определённым электрическим потенциалом мембраны. Можно сказать, что у них двойное управление.
Пример: NMDA-рецепторно-канальный комплекс, имеющий сложную систему управления, включающую в себя 8 рецепторных участков-сайтов, с которыми могут связываться различные лиганды.
6. Опосредованно -управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер-управляемые, управляемые метаботропными рецепторами). Они открываются и закрываются не под действием прямых внешних сигналов, а вследствие опосредованного воздействия на них внутриклеточных вторичных мессенджеров (ионов кальция Са2+, цАМФ, цГМФ, ИФ3, диацилглицерола). Основной механизм такого управления - фосфорилирование ионного канала с внутренней стороны мембраны.
Опосредованное вторичными мессенджерами управление является не прямым, а вторичным. Оно зависит не только от внешнего воздействующего сигнала, но и от наличия, концентрации и активности вторичных мессенджеров. Пусковым сигналом к началу этого процесса может служить воздействие на так называемый метаботропный рецептор, не относящийся к структуре самого управляемого ионного канала и расположенный на мембране где-то отдельно от него. Воздействие на метаботропный рецептор приводит к повышению в клетке концентрации вторичных посредников-мессенджеров. Это ионы Са2+, цАМФ, цГМФ, ИФ3, диацилглицерол. Они активируют соответствующие ферменты-протеинкиназы: А-киназы (цАМФ-зависимые), G-киназы (цГМФ-зависимые), В-киназы (кальций-кальмодулин-зависимые) или С-киназы (кальций-фосфолипид-зависимые). В свою очередь, активированные киназы фосфорилируют ИК мембраны изнутри клетки, т.е. присоединяют к ним фосфаты. В результате этого канал может перейди надолго в новое состояние (открытое или, наоборот, закрытое). После срезания фосфатов ферментом фосфатазой канал возвращается к своему прежнему состоянию. В некоторых случаях такой вторичный мессенджер, как G-белок в виде своей активной субъединицы бета-гамма может сам присоединиться к ионному каналу и поменять его состояние. Так, например, могут открываться (активироваться) калиевые каналы при раздражении ацетилхолином мускариновых рецепторов, связанных с G-белком.
Примеры: Са2+-активируемые хлорные каналы, кальций-активируемые калиевые каналы, цГМФ-активируемые натриевые каналы палочек сетчатки глаза.
Вот, например, опосредованные ион-управляемые кальций-активируемые хлорные каналы являются одним из основных компонентов системы эпителиальной секреции, сенсорной трансдукции, регулирования нейронной и сердечной возбудимости у животных. В клетках растений кальций-активируемые хлорные каналы ответственны за состояние тургора клетки. При повышении концентрации ионов кальция внутри клетки эти каналы открываются и начинают пропускать ионы хлора.
К опосредованно-управляемым (мессенджер-управляемым) ионным каналам формально можно отнести также все каналы, которые управляются "изнутри" с помощью посредников - вторичных мессенджеров. Такой способ управления чаще всего является дополнительным по отношению к "внешнему управлению", и получается, что к мессенджер-управляемым каналам относится большинство каналов из других групп нашей функциональной классификации. Например, все те ИК, которые могут подвергаться фосфорилированию.
7. Актин-управляемые (актин-регулируемые, actin-regulated, actin-gated channels). Они открываются и закрываются за счёт разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков.
В электроневозбудимых клетках активация и инактивация актин-управляемых потенциал-независимых натриевых каналов контролируется процессами разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков. Актиновые элементы цитоскелета, по-видимому, представляют важнейшую часть потенциал-независимого воротного механизма, управляющего открыванием и закрыванием каналов. Именно сборка микрофиламентов на цитоплазматической стороне мембраны приводит к инактивации таких каналов.
8. Коннексоны (двойные поры). Образуют в мембранах контактирующих клеток сквозные непрерывные каналы через две мембраны сразу в зоне щелевых контактов для взаимного обмена веществами между этими клетками. Через коннексоны передаются электрические сигналы, аминокислоты и небольшие молекулы управляющих веществ: цАМФ, InsP3, аденозин, АДФ и АТФ. Они состоят из 6 белковых субъединиц (коннексинов), живущих всего несколько часов. Коннексины - это политопные интегральные мембранные белки 4 раза прошивающие мембрану, имеющие две внеклеточные петли (EL-1 и EL-2), цитоплазматическую петлю (CL) с N-концом (AT) и C-концом (CT), вдающимися в цитоплазму. Через коннексоны соединяется внутренняя среда соседствующих клеток.
Коннексоны являются "неспецифически-управляемыми" каналами. Их состояние регулируется pH, электрическим потенциалом, ионами Са2+, фосфорилированием и другими факторами.
Коннексоны найдены практически во всех видах клеток.
9. «Энерго-зависимые транспортёры» (ионные насосы, ионные помпы, ионные обменники, транспортёры). Это особая группа динамичных пор, проводящих ионы через мембрану, которые формально не относятся к ИК. Их деятельность обеспечивается энергией расщепления АТФ. Они представлены мембранными ферментными белками АТФазами, которые активно протаскивают через себя ионы, используя для этого энергию расщепления АТФ, и обеспечивают активный транспорт ионов через мембрану даже против их градиента концентрации.
Примеры: натрий-калиевый насос, протонный насос, кальциевый насос.
Смотри: 1_2 Транспортные механизмы мембраны. Работа такого транспортёра-насоса описана тут: Механизм натрий-калиевого насоса
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 850; Нарушение авторских прав?; Мы поможем в написании вашей работы!