КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Волновая и квантовая оптика. 1 страница
|
|
|
|
Нейронная сеть. Искусственная нейронная сеть представляет собой совокупность искусственных нейронов, определённым образом соединённых друг с другом и с внешней средой. С инженерной точки зрения такая сеть является распараллеленной динамической системой с топологией направленного графа, которая может выполнять переработку информации посредством изменения своего состояния в ответ на постоянный или импульсный входной сигнал.
План
I. Система органического мира как отражение его исторического развития.
II. Гипотезы возникновения жизни на Земле.
III. Периодизация исторического развития жизни.
IV. Первичные прокариотные экосистемы и эволюция прокариот.
V. Появление эукариот и многоклеточных организмов, их эволюция на протяжении протерозойской эры.
VI. Развитие жизни в палеозойскую эру. Становление современных границ биосферы.
VII. Основные эволюционные события мезозойской эры.
VIII. Развитие жизни в кайнозойскую эру. Человек и его деятельность как фактор эволюции.
I. В составе современной биосферы насчитывают более 3 млн видов живых существ, из них животных - около 2 млн, растений - приблизительно 600 тыс., остальные - это грибы, прокариоты и вирусы. Жизнь на Земле существует около 3,8 млрд. лет. Ученые подсчитали, что в виде ископаемых остатков сохранилось не более 0.1-1.0% вымерших видов, которых на сегодняшний день известно около миллиона. Поэтому реальное число видов, когда-либо живших на нашей планете, составляет от 100 миллионов до миллиарда.
Вам уже известно, что ученые-систематики определяют принадлежность организмов к тому или иному виду. Различные виды объединяют в группы (таксоны) высших рангов (категорий), каждая из которых имеет индивидуальное научное название на латинском языке.
Основные систематические категории (в порядке возрастания рангов):
1) вид,
2) род,
2) семейство,
4) отряд (в ботанике— порядок),
5) класс,
6) тип (в ботанике - отдел)
7) царство.
Любой вид, независимо от того, существует он ныне или вымер, необходимо классифицировать, то есть отнести к каждой из указанных категорий. Например, собака домашняя имеет следующее систематическое положение:
Вид Собака домашняя — Canis familiaris
Род Собака - Canis
Семейство Собачьи – Canidae
Отряд Хищные - Carnivora
Класс Млекопитающие – Mammalia
Тип Хордовые – Chordata
Царство Животные – Animalia
Кроме основных, в систематике некоторых групп организмов употребляют еще и дополнительные (необязательные) систематические категории. Наиболее употребляемые из них - подвид, подрод, подсемейство, надсемейство, подкласс, надкласс и другие. Например, собака принадлежит к подтипу Позвоночные (Vertebrata) и подцарству Многоклеточные (Меtazoa).
Системы живых организмов бывают искусственные и естественные. Основной критерий для построения искусственной (формальной) системы - это степень подобия, классифицируемых объектов, без какого-либо учета степени их исторического родства. Такие системы создают из-за недостатка сведений о той либо иной группе, в частности, об особенностях строения и онтогенеза ее представителей, ископаемых формах. Например, длительное время в искусственный тип Черви объединяли плоских, круглых, кольчатых червей и некоторых других животных. При построении естественных (филогенетических) систем используют следующие принципы: все современные виды—это потомки ископаемых форм, что обусловлено непрерывностью жизни; вследствие дивергенции каждая систематическая группа имеет общего предка. Таким образом, степень подобия видов является следствием общности их происхождения и она тем меньше, чем дальше сравниваемые виды разошлись в процессе исторического развития в результате дивергенции.
II. Проблема происхождения и сущности жизни издавна волновала не только ученых и мыслителей, но и широкие слои населения. Одна из основных догм (догма - утверждение, требующее веры без каких-либо доказательств) всех религий - сотворение жизни нематериальной божественной силой. Эти догмы, например, изложены в Библии. Вера в Бога или атеизм являются личным делом каждого человека.
Многие знаменитые биологи были глубоко верующими людьми (К. Линней, Ч. Дарвин. И.П. Павлов), а Г. Мендель - даже настоятелем монастыря; другие (Ж.-Б. Ламарк, Э. Геккель. И.И. Шмальгаузен) - атеистами, что никак не отразилось на их научных достижениях. Мы рассмотрим только научные гипотезы происхождения жизни.
Гипотезы происхождения жизни подразделяют на абиогенные (живые существа образовались из неживой материи) и биогенные (живые существа могут образовываться только от других живых существ).
Еще древнегреческие ученые (Демокрит, Эпикур, Платон, Аристотель и др.) считали, что живые организмы возникают из ила, навоза, воды, почвы и прочего. На протяжении ХVII-ХVIII столетий такие представления постепенно были отброшены вследствие изучения физиологии размножения многоклеточных организмов. Так, К. Линней в основу разработанной им научной систематики положил принцип «всякий организм происходит от себе подобного». Ученые окончательно отказались от теории абиогенеза во второй половине XIX столетия после открытия известного положения клеточной теории о том. что всякая клетка образуется исключительно вследствие деления материнской клетки. В это же время знаменитый французский биолог Л. Пастер в серии опытов доказал невозможность самозарождения микроорганизмов на примере дрожжей и бактерий.
С развитием геологических знаний об историческом развитии оболочек Земли и молекулярной биологии гипотезы абиогенеза вновь получили развитие в XX столетии. Их особенность состоит в том, что, исключая возможность самозарождения жизни в наше время, ученые считают возможным ее возникновение из химических соединений в прошлом. Впервые подобные взгляды высказал Ж.-Б. Ламарк еще в 1820 г., позже их поддержали Э. Геккель и К.А. Тимирязев. Они считали, что в первичном океане в результате определенных химических процессов возникли вначале органические вещества, затем — доклеточные формы жизни, которые постепенно превратились в клеточные организмы.
В 20-х годах XX столетия русский ученый А.И.Опарин и английский - Д.Холдейн сформулировали биохимическую гипотезу происхождения жизни. Геологи установили, что литосфера, гидросфера и атмосфера сформировались около 4 млрд лет тому назад. Согласно биохимической гипотезе, биологической эволюции предшествовала химическая, которая длилась несколько сотен миллионов лет до появления жизни. Первичная атмосфере состояла из углекислого газа, метана, аммиака, оксидов серы, сероводорода и водяного пара. Озонового экрана не было, и на поверхность Земли падал поток космического и ультрафиолетового солнечного излучения. Вследствие повышенной вулканической активности в воды Мирового океана и атмосферу из земных недр попадали разнообразные химические соединения.
Подобные условия ученые неоднократно создавали в лабораториях. Эксперименты показали, что в водном растворе неорганических веществ, близком по составу к морской воде, в условиях ионизирующего излучения и пропускания через раствор электрических разрядов (аналог молнии) образовывались некоторые органические соединения - нуклеотиды, аминокислоты, пептидные цепочки, моносахариды и другие. Они образовывали скопления, отграниченные от окружающего водного раствора поверхностью раздела — коацерватные капли, способные существовать довольно длительное время.
Ничего, что напоминало бы живые существа, за 70 лет подобных опытов таким образом получить не удалось. Несмотря на это, авторы биохимической гипотезы утверждают, что коацерватные капли каким-то образом превратились в гипотетические «доклеточные» живые организмы, от которых затем произошли прокариоты. Таким образом, абиогенные гипотезы происхождения жизни до сих пор остаются недоказанными.
Современные биогенные взгляды имеют название гипотезы панспермии. Впервые в начале XX столетия ее сформулировал знаменитый шведский физик С. Аррениус, а развил В.И.Вернадский.
Известно, что споры прокариот могут без потери способности к жизнедеятельности выдерживать длительное пребывание в вакууме при температурах, близких к абсолютному нулю (-273°С), радиационное и ультрафиолетовое облучение, то есть условия открытого космоса. Они легко попадают в верхние слои атмосферы планеты с потоками воздуха и оттуда благодаря мизерной массе могут легко выходить в открытый космос и, наоборот, попадать в атмосферу из межпланетного пространства.
С. Аррениус подсчитал, что давление света оказывает заметное механическое воздействие на частицы диаметром около 0,015 мм, перемещая их. Именно такие размеры имеют споры большинства бактерий. Спора, разгоняясь под действием давления солнечных лучей, за 20 суток может преодолеть расстояние между орбитами Земли и Марса, а за 80 - достичь орбиты Юпитера. Также предполагают, что споры могут переноситься метеоритными телами. Согласно гипотезе панспермии, в космосе присутствуют споры прокариот, которые непрерывно попадают на планеты. Если на планете условия благоприятны, то из спор выходят активные формы различных видов, образующие первичные прокариотные биогеоценозы. Последующая эволюция таких видов-первопоселенцев происходит в различных направлениях соответственно изменениям условий окружающей среды на той или иной планете.
III. В пелеонтологии (науке об ископаемых организмах) и филогенетике (науке о путях исторического развития отдельных групп организмов и жизни в целом) ключевое место занимает разделение истории Земли отдельные этапы. Для этого применяются стратиграфические (геохронологические) шкалы. Первая такая шкала, как отражение естественных этапов геологической истории Земли, была утверждена на VIII сессии Международного геологического конгресса в 1900 г. С тех пор эта шкала постоянно модифицируется. В 1960-е годы шкала получила абсолютную (в годах) датировку изотопными методами измерения возраста пород. Из-за постоянного видоизменения ныне существует множество несколько различающийся шкал, хотя с целью стандартизации на международном и государственном уровне в каждый момент времени закреплена только одна. Далее мы принимаем деление на стратоны (временные отрезки) трех рангов (в порядке убывания: 1) эра; 2) период; 3) эпоха. Но будьте готовы в других источниках встретить иные варианты стратиграфической шкалы.
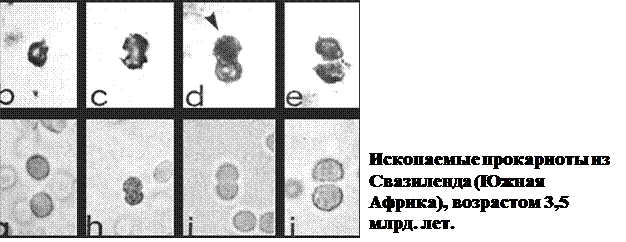
IV. Первые остатки живых организмов найдены в осадочных океанических породах возрастом приблизительно 3,5 млрд. лет, которые сформировались в архейскую эру (началась около 4,5 млрд., окончилась - 2,5 млрд. лет тому назад). Это были прокариоты, представленные остатками карбонатных оболочек колоний цианобактерий и клеточными стенками бактерий разной формы (округлые, палочковидные, нитчатые).
Первичные бактериальные биогеоценозы резко изменили геохимические условия на протяжении архейской эры, при этом изменялись сами. Считают, что первые осадочные породы, имеющие возраст около 3,8 млрд лет; в значительной степени являются следствием деятельности железобактерий (железная руда), зеленых и пурпурных серобактерий (отложения серы, возможно, нефти и природного газа). Благодаря фотосинтезирующей деятельности цианобактерий, в атмосфере постепенно накопилось значительное количество кислорода и образовался озоновый слой. Это привело к возникновению более энергетически выгодного кислородного дыхания и соответственно аэробных прокариот. В конце архейской эры бактерии в морях и океанах полностью осуществляли те же биогенные геохимические процессы, что и сейчас - биогенную миграцию различных химических элементов и превращение энергии в цепях питания.
Эволюционных изменений в строении прокариот практически не происходило: все их ископаемые формы не отличаются от современных. Это объясняют двумя причинами. Во-первых, среди прокариот нет многоклеточных организмов, т.е. они неспособны к дифференциации клеток и усложнению организации. Во-вторых, из-за простого строения генома (единственная кольцевая молекула ДНК) у них легко осуществляется горизонтальный (между различными видами) перенос наследственной информации (вспомните особенности полового процесса прокариот).
У эукариот подобные формы изменения наследственности из-за сложности генома, редки и не являются существенными факторами эволюции.
В прокариотных экосистемах отсутствуют цепи выедания из-за неспособности бактерий к фагоцитозу. Поэтому цепи питания прокариотных биогеоценозов мало разветвлены, а видовое разнообразие в них незначительно.
V. Протерозойская эра началась 2,5 млрд, окончилась - около 0,6 млрд лет тому назад. В ее первой половине прокариотные экосистемы освоили весь Мировой океан. Около 2 млрд. лет тому назад появились первые одноклеточные снабженные жгутиками эукариоты, которые затем дивергировали на растения (водоросли), животных (простейшие) и грибы.
Существует несколько гипотез происхождения эукариот, из которых в наше время наиболее популярна симбиотическая. Ее сторонники считают, что двумембранные органеллы (митохондрии и пластиды), имеющие свой геном и способные к размножению делением, — потомки симбиотических прокариот, утративших способность к существованию вне клетки-хозяина. Так совместное существование нескольких видов прокариот в конце концов привело к появлению эукариотических клеток. Как способ достижения биологического прогресса, для многих эукариот характерно усложнение организации в процессе исторического развития. Уже у одноклеточных (водоросли, инфузории и пр.) клетки устроены довольно сложно. Появление многоклеточных организмов - еще одно свидетельство способности эукариот к усложнению организации. Большинство ученых считает, что многоклеточные организмы произошли от колониальных вследствие дифференциации клеток последних в конце протерозойской эры.
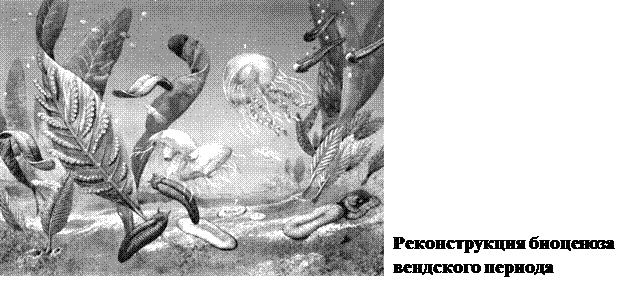 Вендский период - последний период протерозойской эры - длился около 80 млн лет. В это время на мелководьях морей образовались биогеоценозы, основными продуцентами в которых были цианобактерии и зеленые водоросли. Наиболее распространенными животными были разнообразные кишечнополостные, представленные как полипами, так и медузами. Некоторые из них достигали метра в диаметре. От таких форм с лучевой симметрией в середине периода произошли ползающие и плавающие двустороннесимметричные виды. Среди них были несегментированные и сегментированные организмы; у некоторых из последних на каждом сегменте была пара конечностей. В конце периода в связи с биосферным кризисом, вызванным оледенением, вендские экосистемы были разрушены и большинство составляющих их видов вымерло.
Вендский период - последний период протерозойской эры - длился около 80 млн лет. В это время на мелководьях морей образовались биогеоценозы, основными продуцентами в которых были цианобактерии и зеленые водоросли. Наиболее распространенными животными были разнообразные кишечнополостные, представленные как полипами, так и медузами. Некоторые из них достигали метра в диаметре. От таких форм с лучевой симметрией в середине периода произошли ползающие и плавающие двустороннесимметричные виды. Среди них были несегментированные и сегментированные организмы; у некоторых из последних на каждом сегменте была пара конечностей. В конце периода в связи с биосферным кризисом, вызванным оледенением, вендские экосистемы были разрушены и большинство составляющих их видов вымерло.
VI. Палеозойская эра началась около 600 млн. и окончилась 240 млн. лет тому назад. На протяжении этого времени жизнь освоила сушу и биосфера достигла современных пределов.
1. Кембрийский период (600-500 млн. лет тому назад) отличался преимущественно теплым климатом. Жизнь концентрировалась в основном в неглубоких морях с температурой воды +20...25° С.
С этого периода известны остатки зеленых и красных водорослей, а также одноклеточных животных - фораминифер. Появляются губки — особый тип неподвижно прикрепленных многоклеточных животных, состоящих из различных типов клеток, не образующих тканей, и питающихся путем фильтрации. Кембрийские губки имели массивный скелет из углекислого кальция. Их колонии образовывали рифы. Они принадлежали к отдельному классу, один из представителей которого существует и ныне. Кишечнополостные кембрийского периода были менее разнообразны, чем вендского.
Из кембрийских отложений известны хорошо сохранившиеся остатки многощетинковых червей и членистоногих - ракообразные и особые, ныне вымершие, формы - трилобиты. Появляются представители всех известных классов моллюсков - брюхоногие, двустворчатые, головоногие. Последние имели внешнюю раковину.
В этот период жили представители 8 классов типа Иглокожие (сейчас насчитывают только 6 классов этих животных). Это исключительно морские обитатели с лучевой, реже - двусторонней симметрией. Они обладают скелетом из углекислого кальция, вторичной полостью тела и особой водно-сосудистой системой, служащей для передвижения, захвата пищи и дыхания. Считают, что иглокожие произошли от общих с хордовыми предков.
Появляются первые хордовые, которые в общем напоминали современного ланцетника, а в конце периода - и представители позвоночных (бесчелюстные).

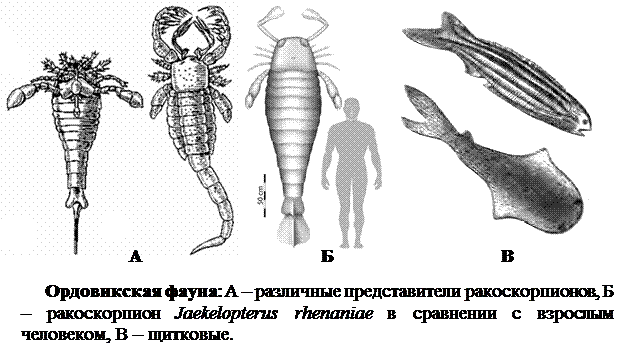
2. Ордовикский период окончился около 440 млн. лет тому назад. В морях достигли большого видового разнообразия головоногие, конусовидные раковины некоторых из них достигали 9 м в длину. Известно около 10 классов иглокожих, среди которых также были гигантские формы - морские лилии длиной до 20 м. Появляются разные коралловые полипы, которые стали основными рифообразователями. Жизнь освоила пресные водоемы, где росли зеленые водоросли, обитали разнообразные ракообразные и самые крупные за всю историю Земли членистоногие - ракоскорпионы. Эти хищники достигали 2 м в длину.
В это время произошел расцвет бесчелюстных позвоночных - щитковых. Они имели обтекаемую рыбообразную форму тела, непарные, а иногда и парные, плавники и хрящевой скелет. Большинство видов снаружи защищали щитки из костной ткани, которые часто были слиты в сплошной панцирь. У них не было жаберных дуг и челюстей; органами дыхания служили жаберные мешки, как у современных миног. Длина тела этих животных составляла от нескольких сантиметров до метра. Щитковые, в основном, питались мелким планктоном и остатками организмов.
3. Силурийский период (440-400 млн. лет тому назад) характеризуется наличием мелководных (до 10 м глубины) теплых морей с пониженной соленостью. В них появились первые челюстные позвоночные: особые ископаемые классы рыб.
Силурийские рыбы имели внутренний скелет из хрящевой ткани, у них не было жаберных крышек и плавательного пузыря. Колючезубые достигали не более 30 см длины. Для них характерно наличие до 8 пар плавников. Их тело покрывала костная чешуя. Панцирные рыбы были покрыты костными пластинками, часто слитыми в сплошной панцирь. Некоторые из них достигали 6 м длины. Среди силурийских рыб были придонные и плавающие в толще воды формы. Большинство из них, как полагают ученые, были хищниками.
В прибрежных частях континентальных водоемов из-за колебаний уровня воды образовался слой ила. Он стал основой первичных почв, на которых образовались наземные биогеоценозы, состоящие в основном из высших споровых растений из отделов Риниофиты и Плаунообразные. Из животных здесь обитали почвенные виды (малощетинковые черви и др.) и питающиеся остатками растений двупарноногие многоножки. Это особый класс членистоногих, представители которых и сейчас широко распространены на Земле. Их тело состоит из головы и многосегментного туловища; каждый сегмент несет 2 пары ног. Известны и первые наземные хищники - скорпионы.

4. Девонского период окончился около 350 млн лет тому назад. В это время процессы горообразования чередовались с опусканием суши, что вызывало частые биоценотические кризисы. Поэтому темпы эволюции были сравнительно высокие.
В начале периода вымерла большая часть трилобитов, а в его конце - риниофиты, панцирные рыбы и щитковые. На пониженных увлажненных частях суши образовались леса, состоящие из деревянистых высших споровых - плаунов, хвощей и папоротников. Появились первые голосеменные из класса семенных папоротников. Эти растения по внешнему виду напоминали древовидные папоротники, но размножались семенами. Из беспозвоночных сушу освоили пауки и клещи.
В морях появились хрящевые рыбы (акулы и некоторые другие). В пресных водоемах сформировались костные рыбы, в основном кистеперые и двоякодышащие, имевшие значительное видовое разнообразие, а также первые лучеперые. Считают, что плавательный пузырь костных рыб вначале служил как легкое для дополнительного дыхания атмосферным кислородом, так как в воде пресных водоемов, где перегнивали остатки древесных растений, растворенного кислорода было недостаточно.
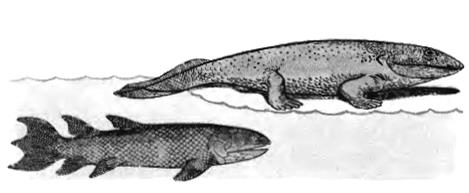
Мускулистые парные плавники кистеперых рыб, вероятно, служили им не столько для плавания, сколько для ползания по дну водоемов между стволами упавших деревьев. План строения этих плавников оказался удачным для превращения их в пригодные для передвижения по суше конечности. Потомки некоторых кистеперых перешли к жизни на суше, где нашли богатую кормовую базу (различных беспозвоночных), но их размножение и развитие было еще связано с водой. Так произошли земноводные. В связи с дыханием атмосферным кислородом у них резко возросло количество гемоглобина, поэтому основным кроветворным органом у наземных позвоночных становится красный костный мозг (у рыб - только селезенка). Девонские земноводные имели самые разнообразные размеры (от нескольких сантиметров до 3-4 м длины) и форму тела.
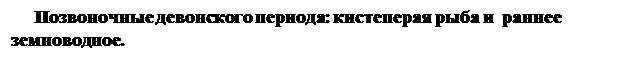
5. Каменноугольный период окончился около 280 млн. лет тому назад. Он был одним из самых теплых в истории Земли. На суше было много увлажненных низин, где произрастали леса из разнообразных высших споровых и голосеменных. Попадая в заболоченную почву, стволы упавших деревьев в условиях отсутствия кислорода не перегнивали, а заиливались, покрывались слоями песка и глины и в конце концов оказывались на значительных глубинах под землей, где в условиях высокого давления превратились в каменный уголь (отсюда и название периода).
В это время появились хвойные растения, размножение которых не было связано с наличием воды. Они образовали основу биогеоценозов среднеувлажненных и засушливых местностей, и в конце периода вся суша была освоена живыми существами, то есть биосфера достигла современных границ. В этот период появляются мохообразные.
Вслед за растениями быстрыми темпами осваивали сушу животные. В начале периода, верочтно, от каких-то пресноводных ракообразных произошли вначале бескрылые, а затем - крылатые насекомые. Насекомые стали первыми летающими животными. В конце периода уже существовало 15 отрядов насекомых. Интересно, что некоторые виды каменноугольных стрекоз имели размах крыльев до метра. На суше появились легочные брюхоногие моллюски.
В середине периода некоторые земноводные приспособились к размножению на суше. Этому способствовало внутреннее оплодотворение, образование богатых запасными веществами и покрытых толстыми водонепроницаемыми оболочками яиц и прямое развитие. У одних из них кожные железы исчезли, а сама кожа стала очень толстой (приспособление к сохранению воды). Они дали начало первым пресмыкающимся. У других видов эти железы сохранились как приспособления к терморегуляции путем испарения воды через кожу. Такие виды стали предками последовательного ряда форм, которые в конце концов привели к млекопитающим.

 6. Пермский период (280-240 млн. лет тому назад) характеризовался снижением уровня Мирового океана. На протяжении этого периода произошло несколько оледенений значительной части суши и, соответственно, биосферных кризисов, а также связанные с ними вымирания одних и появление других систематических групп организмов.
6. Пермский период (280-240 млн. лет тому назад) характеризовался снижением уровня Мирового океана. На протяжении этого периода произошло несколько оледенений значительной части суши и, соответственно, биосферных кризисов, а также связанные с ними вымирания одних и появление других систематических групп организмов.
Основу наземных биогеоценозов составляли хвойные и некоторые другие голосеменные, а древовидные высшие споровые почти полностью исчезли. Происходило дальнейшее развитие насекомых: к концу периода их насчитывалось 30 отрядов, в том числе прямокрылые, жесткокрылые, чешуйчатокрылые и перепончатокрылые. Значительно возросло разнообразие пресмыкающихся: появились черепахи, ящерицы и некоторые другие формы.
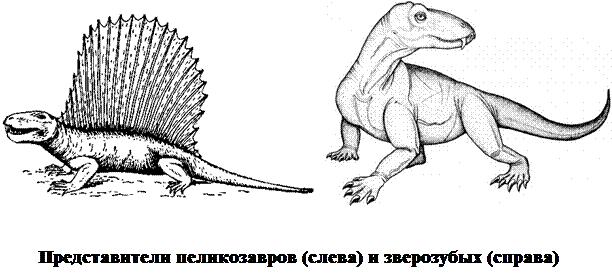
Линия развития наземных позвоночных, ведущая к млекопитающим, в этот период вначале была представлена пеликозаврами - хищниками до 4 м длиной с хорошо развитыми клыками. На смену им пришли зверозубые, среди которых были как мелкие насекомоядные виды, так и крупные хищники длиной до 7 м. Кроме резцов и клыков у них были развиты также и коренные зубы, приспособленные к пережевыванию пищи.
К концу периода в морях полностью вымерли трилобиты, колючезубые рыбы и многие другие группы, а в пресных водоемах - значительная часть двоякодышащих, кистеперых рыб и земноводных. Итак, в биосфере произошли изменения, которые подготовили господство голосеменных и пресмыкающихся на протяжении следующей мезозойской эры.
VII. Мезозойская эра началась 240 и окончилась 63 млн лет тому назад. В ней выделяют триасовый, юрский и меловой периоды.
1. Триасовый период окончился приблизительно 185 млн. лет тому назад. Его климатические условия напоминали пермский. В морях увеличилось видовое разнообразие хрящевых и костных рыб, последние распространились и в соленых водоемах. С этого периода известны представители всех современных и нескольких ископаемых отрядов насекомых.
Появились два отряда наземных пресмыкающихся, известных под названием динозавры. Они имели средние или большие (от одного до 30 м длины) размеры, передвигались на четырех или двух задних конечностях, направленных вниз, в отличие от современных пресмыкающихся. Существовало несколько групп морских пресмыкающихся. От насекомоядных зверозубых небольших размеров во второй половине периода произошли первые млекопитающие. Они были покрыты шерстью и внешне напоминали современных землероек, их длина была в пределах 5 - 15 см. Неизвестно, были ли они живородящими или откладывали яйца подобно современным первозверям.
Со средины периода известны крокодилы, которые, в отличие от современных, были очень подвижными наземными хищниками с удлиненными приспособленными к бегу конечностями.
Триасовый период окончился биосферным кризисом, связанным с подъемом уровня океана и наступившим вследствие этого потеплением климата. Это сопровождалось вымиранием многих групп — древних земноводных, зверозубых и других.
2. Юрский период (185-130 млн. лет тому назад) характеризовался умеренным климатом. В это время существовало много мелководных морей. В них достигли значительного видового разнообразия головоногие моллюски, которые в общем напоминали современных кальмаров, но имели прямую внешнюю раковину — белемниты. Остатки этих раковин, которые часто находят в песчаных породах, известны в народе под названием «чертовы пальцы». Появились и распространились в пресных и соленых водоемах диатомовые водоросли.
Крокодилы перешли к обитанию в водной среде и приняли современный облик. Интересно, что наряду с пресноводными существовали и морские виды, некоторые из последних достигали 15 м длины. Были распространены водные хищные пресмыкающиеся — плезиозавры и ихтиозавры.
Плезиозавры были от 2 до 20 м длиной. Голова находилась на длинной гибкой шее (число шейных позвонков достигало 80); рот был вооружен острыми зубами. Туловище было бочонкообразное; обе пары конечностей превратились в ласты. Они занимали в биоценозах место современных ластоногих и размножались на суше.
Ихтиозавры (длиной от одного до 15 м) имели рыбообразную форму. У них была короткая шея, передние и задние конечности преобразованы в ласты, а хвост - в вертикальный плавник. Подобно современным зубатым китам, они никогда не выходили на сушу и были живородящими (найден скелет самки со сформированным детенышем внутри брюшной полости).
Птерозавры, или летающие ящеры, появились в конце триасового периода, но наибольшего видового разнообразия достигли в юрский и меловой периоды. Подобно современным птицам, у них были полые кости, киль грудины, облегченный череп с тонкими костями, на челюстях были мелкие зубы или роговые чехлы, напоминающие клюв. Как и у современных рукокрылых, крылья птерозавров представляли собой кожистую перепонку, натянутую с боков тела от передних до задних конечностей. Она прикреплялась к очень длинному пятому пальцу (мизинцу) передних конечностей.
Динозавры были очень разнообразны. На четырех конечностях передвигались преимущественно растительноядные формы. Из них наиболее известны диплодок (длиной около 30 м) и похожий на него, но более массивный бронтозавр (с массой около 80 т). Это были самые большие сухопутные животные за всю историю Земли. На территории современной Северной Америки обитал стегозавр, на спине которого располагалось два ряда параллельных вертикальных костных пластин. Предполагают, что через них организм рассеивал избыток тепла.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 1107; Нарушение авторских прав?; Мы поможем в написании вашей работы!