КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Посттранскрипционные процессы
|
|
|
|
Процессинг - это превращение первичного транскриптата в зрелую и-РНК, выполняющей роль матрицы при трансляции.
Стадии процессинга:
1. Отщепление концевых участков первичного транскриптата.
2. Формирование на 5' конце колпачка.
3. Формирование на 3' конце полиоадениловой последовательности нуклеотидов.
4. Метилирование некоторых азотистых оснований в транскрип-тате, стабилизирующее молекулу РНК.
5. Вырезание неинформативных участков, соответствующих ин-тронам ДНК и сшивание (сплайсинг) участков, соответствующих экзонам.
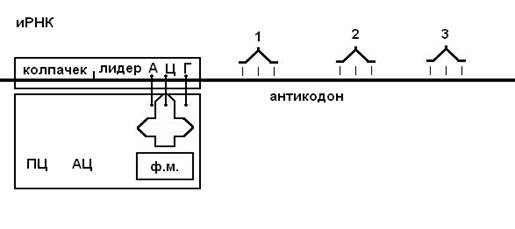
В результате процессинга у эукариот образуется зрелая и-РНК, имеющая следующее строение:
рис. 2 :

Колпачок - последовательность нуклеотидов с метилированными основаниями, которая обеспечивает узнавание малых субъединиц рибосом. Лидер - вводная последовательность нуклеотидов, компли-ментарная последовательности в молекуле р-РНК малой субъединицы рибосомы, которая обеспечивает прикрепление и-РНК к малой субъединице.
Стартовый кодон - триплет нуклеотидов, кодирующий в большинстве случаев аминокислоту формилметионин (АЦГ).
Кодирующая часть - последовательность кодонов, шифрующих определенную последовательность аминокислот в полипептидной цепи.
Трейлер - концевая часть молекулы и-РНК, включающая нон-сенс-кодон или поли-А-последовательность (А-А-А-А).
III. Трансляция.
Трансляция - механизм, с помощью которого последовательность триплетов оснований в молекулах и-РНК переводится в специфическую последовательность аминокислот в полипептидной цепи. Этот процесс происходит на рибосомах.
Основные фазы трансляции:
А. Инициация.
В. Элонгация.
С. Терминация.
В фазу инициации происходят следующие процессы:
А. С помощью колпачка и-РНК находят в цитоплазме малую субъединицу рибосомы.
В. С помощью лидерной последовательности устанавливается связь с с комплиментарным участком р-РНК и р-РНК присоединяется к малой субъединице.
С. К стартовому кодону (АЦГ) присоединяется т-РНК, несущая формилметионин.
D. Малая субъединица ассоциируется с большой субъединицей, в аминоацильном центре (АЦ) которой располагается формил метионин.
Рис. 3
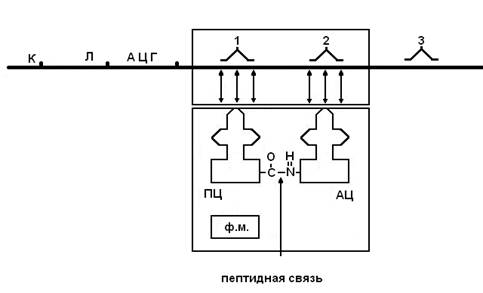
После прохождения через рибосому всей кодирующей части и-РНК на рибосоме собирается пептидная цепь с определенной последовательностью аминокислот. Фаза терминации наступает, когда в контакт с рибосомой приходит концевой участок и-РНК, который включает нонсенс-триплет, не кодирующий никакой аминокислоты. На этом сборка пептидной цепи заканчивается. По мере освобождения 5' конца и-РНК, колпачок может находить новые малые субъединицы рибосом и процесс трансляции может повторно осуществляться на новых рибосомах. Комплекс рибосом, синтезирующих одинаковые полипептидные цепи, называется полирибосомой (полисомой).
ПРИЛОЖЕНИЕ 2. ДОКАЗАТЕЛЬСТВА ГЕНЕТИЧЕСКОЙ РОЛИ НУКЛЕИНОВОЙ КИСЛОТЫ
История изучения нуклеиновых кислот начинается с 1869 г., когда швейцарский химик Ф.Мишер обнаружил в клеточном ядре особые вещества, обладающие свойствами кислот. Он дал им название нуклеиновых кислот (от лат. пис1еиз — ядро). В опытах английского бактериолога Ф.Гриффитса (1928) была продемонстрирована способность пневмококков к трансформации, было выдвинуто предположение о том, что «трансформирующий агент», отождествляемый с «веществом наследственности», находится в ядре. Суть эксперимента Гриффитса заключалась в следующем. При введении мышам непатогенных штаммов пневмококков (рис. 1.) животные не заболевали (Б). При введении патогенных штаммов мыши гибли (А), однако при введении патогенных микробов, убитых нагреванием, мыши оставались здоровыми (В). Гриффитс показал, что при одновременном введении живых непатогенных и убитых патогенных микробов мыши погибали (Г). Гриффитс заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного приобретают наследственно закрепленные свойства патогенности (трансформируются). В последующем было доказано, что трансформация происходит не только в живом организме, но и in vitro, т.е. в пробирке. Следующее замечательное открытие принадлежит О. Эвери, К.МакЛеод и М.МакКарти, которые в 1944 г. точно определили химическую природу «трансформирующего агента» и идентифицировали его как дезоксирибонуклеиновую кислоту. Чистая ДНК, выделенная из клеток патогенного штамма, при добавлении в кулътуру непатогенных клеток
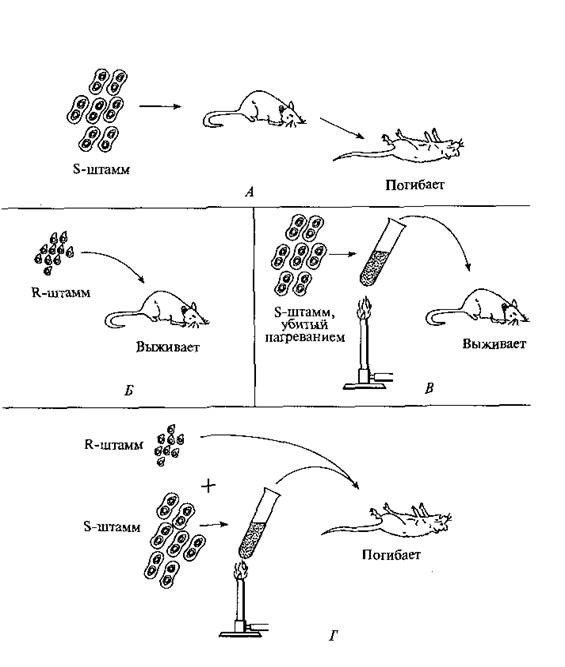
Рис. 1. Схема эксперимента Ф. Гриффитса.
трансформировала последние, придавая им свойства патогенности. Это новое свойство передавалось при дальнейшем размножении. При обработке трансформирующего агента специфическими веществами, разрушающими ДНК, трансформация не осуществлялась. Таким образом, было получено прямое доказательство генетической роли ДНК. Еще одним шагом в доказательстве генетической роли нуклеиновой кислоты является открытие правила эквивалентности, согласно которому в ДНК, выделенных из организмов различных видов, соотношение пуриновых и пиримидиновых оснований всегда одно и то же и составляет 1:1. Второе доказательство роли ДНК в передаче наследственной информации получили Н. Зиндер и Дж. Ледерберг. В 1952 г. они описали явление трансдукции. Они взяли U-образную трубку, между коленами которой находился антибактериальный фильтр. В одно колено поместили бактерии, способные синтезировать триптофан лизогенным бактериофагом, а в другое – триптофан не синтезирующий штамм. Через некоторое время в первом колене погибли все бактерии, а во втором штамм неспособный к синтезу триптофана получил эту способность. То есть произошла трасдукция – способность бактериофагов переносить фрагменты ДНК от одного штамма бактерий к другому и передавать соответствующие свойства. К началу 50-х гг. было получено множество данных (на различных объектах), свидетельствующих об универсальности ДНК как носителя генетической информации. Вирусы, как было сказано ранее, имеют относительно простое строение: они состоят из белковой оболочки, содержащей атомы серы, и заключенной внутри нее молекулы нуклеиновой кислоты, содержащей атомы фосфора. В 1952 г. А. Херши и М. Чейз проводили эксперименты с бактериофагом Т2 — особым видом вируса, убивающим зараженную бактериальную клетку (отсюда и название «бактериофаг», т.е. пожиратель бактерии). Бактериофаг, проникая в кишечную палочку, быстро в ней размножается. Экспериментаторы размножали бактериофаги в клетках Е. coli, которые росли на двух различных средах: на среде, содержащей радиоактивный изотоп серы (35S), и на среде, содержащей радиоактивный изотоп фосфора (32Р). Фаги, которые размножились на клетках, выросших на среде с радиоактивным изотопом серы, включали 35S только в свои белковые оболочки. Фаги, размножившиеся на клетках, которые выросли на среде с радиоактивным фосфором, содержали ДНК, меченную 32Р. Затем полученными бактериофагами заражали клетки Е. соli, выращенные на обычной среде. Через короткое время эти клетки интенсивно встряхивали, чтобы отделить бактериофаги от стенок Е. coli. Затем делали анализ бактерии на наличие радиоактивности. Оказалось, что бактерии, зараженные фагами, выросшими на 35S, не содержали радиоактивной метки, в то время как бактерии, зараженные фагами, размножившимися на 32Р, были радиоактивными. Полученные результаты позволили авторам сделать два принципиальных вывода: 1) в бактериальную клетку проникает только фаговая ДНК, которая, размножаясь в клетке Е. соli, дает начало многочисленному потомству; 2) наследственным материалом является ДНК, которая определяет не только структуру и свойства ДНК потомства, но и свойства фаговых белков. В 1953 г. Дж.Уотсон и Ф. Крик на основании результатов рентгеноструктурного анализа и биохимических данных предложили пространственную модель структуры ДНК, объясняющую все ее свойства. Согласно предложенной модели молекула ДНК состоит из двух комплементарных (соответствующих) нитей. М. Мезельсон и Ф. Сталь доказали полуконсервативный механизм репликации (удвоения) ДНК. Выяснение структуры и функции нуклеиновых кислот позволило понять, каким образом живые организмы воспроизводят себя и как осуществляются кодирование генетической информации, ее хранение и реализация, необходимые для протекания всех жизненных процессов. К настоящему времени существенным образом обогащены знания о структуре и функции ДНК, значительно расширены возможности для исследований. Было обнаружено, что ДНК может как повреждаться, так и восстанавливаться, что молекулы ДНК могут обмениваться друг с другом частями, закручиваться и раскручиваться. Было показано, что ДНК служит матрицей для синтеза РНК, а также сама способна синтезироваться в процессе обратной транскрипции с РНК. ДНК функционирует не только в ядре, но и в митохондриях. В настоящее время исследователи способны определять последовательность нуклеиновых оснований в ДНК и осуществлять ее синтез.
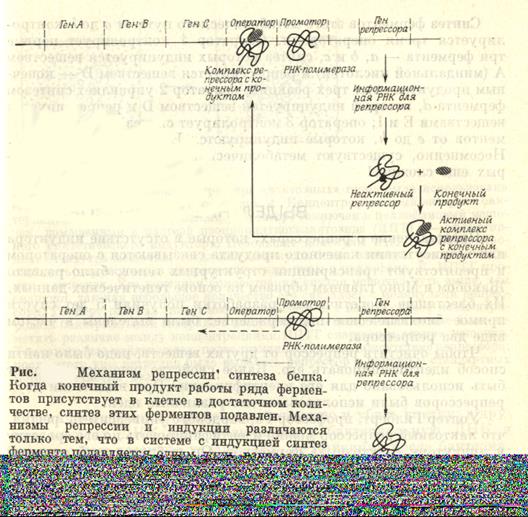
ПРИЛОЖЕНИЕ 3. РЕГУЛЯЦИЯ БИОСИНТЕЗА БЕЛКА БАКТЕРИАЛЬНОЙ КЛЕТКИ
Жакоб и Моно, изучая влияние мутаций на проявление трех бактериальных генов, контролирующих синтез трех ферментов, необходимых для расщепления сахара лактозы, доказали существование двух различных генетических систем. Одна система состоит из структурных генов, каждая из которых определяет структуру фермента или другого белка (три структурных гена лактозной области кодируют последовательность аминокислот в трех ферментах – бета-галоктозидазе, пермеазе, ацетилазе). Другую генетическую систему образуют гены регуляторы(например ген репрессора), ответственные за включение и выключение структурных генов, так что соответствую-щие белки синтезируются или не синтезируются. На первом рисунке показана система регуляции биосинтеза по типу индукции. Отсутствие в клетке лактозы приводит к тому, что белок репрессор соединяется с оператором оперона и синтез ферментов для расщеп-ления лактозы не идет (рис. А). При появлении лактозы белок репрессор соединяется с ней и оператор открыт для синтеза информации о нужных ферментах (рис.Б).
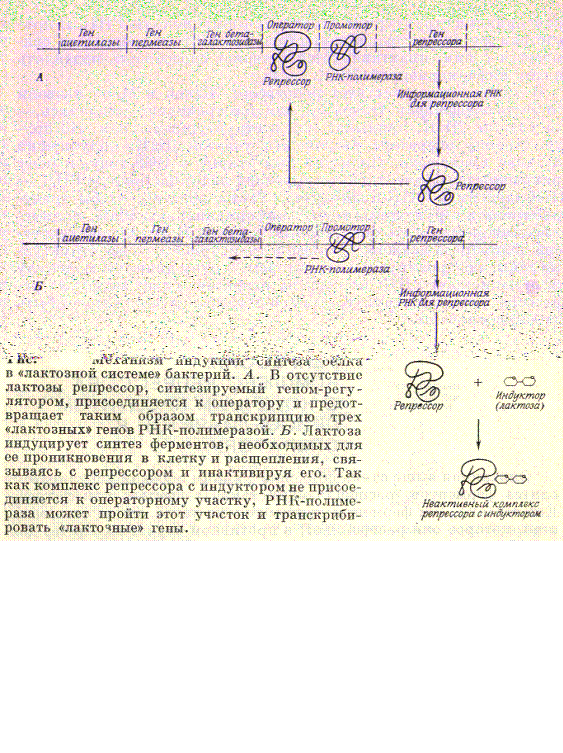 |
Регуляция по типу репрессии. Этот механизм отличается от индукции только одной деталь: белок репрессор неактивен, поэтому он соединяется с продуктом, который должен достичь критической концентрации. В этом случае, активизируется комплекс: продукт + белок репрессор соединяется с оператором и закрывает ферменты, которые отвечают за синтез самого продукта.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 1892; Нарушение авторских прав?; Мы поможем в написании вашей работы!