КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эмбриональное развитие
|
|
|
|
| Дробление |
После оплодотворения зигота начинает делиться. Дроблением называют ряд последовательных митотических делений зиготы, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные, содержащие ядра клетки меньшего размера (рис. 312). В результате дробления образуются клетки, которые называют бластомерами. Важной отличительной особенностью дробления от обычного деления является то, что вновь образовавшиеся бластомеры не увеличиваются в размерах. Это осуществляется путем выпадения интерфазного периода роста между делениями. При этом синтетический период интерфазы начинается в телофазе предшествующего митотического цикла. Таким образом, количество бластомеров постепенно увеличивается, а их общий объем практически не изменяется. Цитоплазма клеток при дроблении делится путем возникновения впячиваний оболочки клетки (борозды дробления).
|
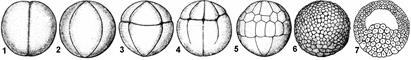 Рис. 312. Дробление яйцеклетки амфибий (лягушка):
1 — двуклеточная стадия; 2 — четырехклеточная стадия; 3 — восьмиклеточная стадия; 4 — переход от восьми- к шестнадцатиклеточной стадии (клетки анимального полюса уже поделились, а клетки вегетативного только начинают дробиться; 5 — более поздняя стадия дробления; 6 — бластула; 7 — бластула в разрезе.
Рис. 312. Дробление яйцеклетки амфибий (лягушка):
1 — двуклеточная стадия; 2 — четырехклеточная стадия; 3 — восьмиклеточная стадия; 4 — переход от восьми- к шестнадцатиклеточной стадии (клетки анимального полюса уже поделились, а клетки вегетативного только начинают дробиться; 5 — более поздняя стадия дробления; 6 — бластула; 7 — бластула в разрезе.
Однако дробление не может происходить бесконечно. Так как каждое деление дробления сопровождается уменьшением размера клетки, постепенно происходит повышение величины ядерно-цитоплазматического отношения, сниженного в период роста ооцита. Наступает момент, когда это отношение достигает значения, типичного для соматических клеток данного вида.
Биологическое значение процесса дробления сводится к следующему:
© благодаря повторяющимся циклам репродукции, происходит размножение генотипа зиготы;
© происходит накопление клеточной массы для дальнейших преобразований, т.е. зародыш из одноклеточного превращается в многоклеточный.
Деление бластомеров бывает синхронным и несинхронным. У большинства видов оно несинхронно с самого начала развития, у других становится таковым уже после первых делений.
Характер дробления определяется, прежде всего, строением яйцеклетки, главным образом, количеством желтка и особенностями его распределения в цитоплазме. В этой связи по способу дробления выделяют два основных типа яиц (рис. 313):
© полностью дробящиеся;
© дробящиеся частично.
| Полное дробление |
Полным дробление называется тогда, когда цитоплазма яйцеклетки полностью разделяется на бластомеры. Оно может быть:
© равномерным, при котором все образовавшиеся бластомеры имеют одинаковые размеры и форму; оно характерно для алецитальных и изолецитальных яйцеклеток;
© неравномерным, при котором образуются неравные по размерам бластомеры; свойственно телолецитальным яйцеклеткам с умеренным содержанием желтка; мелкие бластомеры возникают у анимального полюса, крупные — в области вегетативного полюса зародыша.
|
 Рис. 313. Различные типы дробления:
А — полное; Б — частичное; В — дискоидальное.
Рис. 313. Различные типы дробления:
А — полное; Б — частичное; В — дискоидальное.
| Частичное дробление |
Частичное дробление — тип дробления, при котором цитоплазма яйцеклетки не полностью разделяется на бластомеры. Одним из видов частичного дробления является дискоидальное, при котором дроблению подвергается только лишенный желтка участок цитоплазмы у анимального полюса, где находится ядро. Участок цитоплазмы, подвергшийся дроблению, называется зародышевым диском. Этот тип дробления характерен для резко телолецитальных яиц с большим количеством желтка (рептилии, птицы, рыбы);
| Образование бластулы |
Дробление у представителей разных групп животных имеет свои особенности, однако завершается оно образованием близкой по строению структуры — бластулы.
Бластула — это однослойный зародыш. Она состоит из слоя клеток — бластодермы, ограничивающей полость — бластоцель, или первичную полостью тела. Бластула формируется начиная с ранних этапов дробления, благодаря расхождению бластомеров. Возникающая при этом полость заполняется жидкостью.
| Типы бластул |
Строение бластулы во многом зависит от типа дробления (рис. 314).
© Целобластула (типичная бластула). Образуется при равномерном дроблении. Имеет вид однослойного пузырька с большим бластоцелем (у ланцетника).
© Амфибластула. При дроблении телолецитальных яиц бластодерма построена из бластомеров разного размера: микромеров на анимальном и макромеров на вегетативном полюсах. Бластоцель при этом смещается в сторону анимального полюса (у земноводных).
|
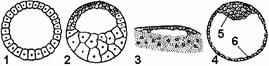 Рис. 314. Типы бластул:
1 — целобластула; 2 — амфибластула; 3 — дискобластула; 4 — бластоциста; 5 — эмбриобласт; 6 — трофобласт.
Рис. 314. Типы бластул:
1 — целобластула; 2 — амфибластула; 3 — дискобластула; 4 — бластоциста; 5 — эмбриобласт; 6 — трофобласт.
© Дискобластула. Образуется при дискоидальном дроблении. Полость бластулы имеет вид узкой щели, находящейся под зародышевым диском (у птиц).
© Бластоциста. Представляет собой однослойный пузырек, заполненный жидкостью, в котором различают эмбриобласт (из него развивается зародыш) и трофобласт, обеспечивающий питание зародыша (у млекопитающих).
| Гаструляция |
После того как сформировалась бластула, начинается новый этап эмбриогенеза — гаструляция (образование зародышевых листков). Для гаструляции характерны интенсивные перемещения отдельных клеток и клеточных масс. Деление клеток при гаструляции отсутствует или выражено очень слабо. В результате гаструляции образуется двухслойный, а затем трехслойный зародыш (у большинства животных) — гаструла (рис. 315). Первоначально образуются наружный (эктодерма) и внутренний (энтодерма). Позже между экто- и энтодермойзакладывается третий зародышевый листок — мезодерма.
Зародышевые листки — это отдельные пласты клеток, занимающие определенное положение в зародыше и дающие начало соответствующим органам и системам органов. Зародышевые листки возникают не только в результате перемещения клеточных масс, но и в результате дифференциации сходных между собой сравнительно однородных клеток бластулы. В процессе гаструляции зародышевые листки занимают положение, соответствующее плану строения взрослого организма. Дифференциация — это процесс появления и нарастания морфологических и функциональных различий между отдельными клетками и частями зародыша.
| Способы гаструляции |
|
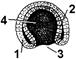 Рис. 315. Гаструла.
1 — эктодерма; 2 — энтодерма; 3 — бластопор;
4 — гастроцель.
Рис. 315. Гаструла.
1 — эктодерма; 2 — энтодерма; 3 — бластопор;
4 — гастроцель.
© Инвагинация. При данном способе один из участков бластодермы начинает впячиваться внутрь бластоцеля (у ланцетника). При этом бластоцель практически полностью вытесняется. Образуется двухслойный мешок, наружная стенка которого является первичной эктодермой, а внутренняя — первичной энтодермой, выстилающей полость первичной
кишки, или гастроцель. Отверстие, при помощи которого полость сообщается с окружающей средой, называется бластопором, или первичным ртом. У представителей разных групп животных судьба бластопора различна. У первичноротых животных он превращается в ротовое отверстие. У вторичноротых бластопор зарастает, и на его месте нередко возникает анальное отверстие, а ротовое отверстие прорывается на противоположном полюсе (переднем конце тела).
|
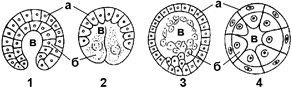 Рис. 316. Типы гаструл:
1 — инвагинационная; 2 — эпиболическая; 3 — иммиграционная; 4 — деламинационная; а — эктодерма; б — энтодерма; в — гастроцель
Рис. 316. Типы гаструл:
1 — инвагинационная; 2 — эпиболическая; 3 — иммиграционная; 4 — деламинационная; а — эктодерма; б — энтодерма; в — гастроцель
© Иммиграция — выселение части клеток бластодермы в полость бластоцеля (у высших позвоночных). Из них образуется энтодерма.
© Деламинация встречается у животных, имеющих бластулу без бластоцеля (у птиц). При таком способе гаструляции клеточные перемещения минимальны или совсем отсутствуют, так как происходит расслоение — наружные клетки бластулы преобразуются в эктодерму, а внутренние формируют энтодерму.
© Эпиболия происходит, когда более мелкие бластомеры анимального полюса дробятся быстрее и обрастают более крупные бластомеры вегетативного полюса, образуя эктодерму (у земноводных). Клетки вегетативного полюса дают начало внутреннему зародышевому листку — энтодерме.
Описанные способы гаструляции редко встречаются в чистом виде и обычно наблюдаются их сочетания (инвагинация с эпиболией у амфибий или деляминация с иммиграцией у иглокожих).
| Образование мезодермы |
Чаще всего клеточный материал мезодермы входит в состав энтодермы. Он впячивается в бластоцель в виде карманообразных выростов, которые затем отшнуровываются.
При образовании мезодермы происходит образование вторичной полости тела, или целома.
| Первичный органогенез |
Процесс формирования органов в эмбриональном развитии называют органогенезом. В построении любого органа участвуют несколько тканей. Поэтому стадия органогенеза является и стадией гистогенеза.
В органогенезе можно выделить две фазы:
© нейруляция — образование комплекса осевых органов (нервная трубка, хорда, кишечная трубка и мезодерма сомитов), в который вовлекается почти весь зародыш;
© построение остальных органов, приобретение различными участками тела типичной для них формы и черт внутренней организации, установление определенных пропорций (пространственно ограниченные процессы).
По теории зародышевых листков Карла Бэра, возникновение органов обусловлено преобразованием того или иного зародышевого листка — экто-, мезо- или энтодермы. Некоторые органы могут иметь смешанное происхождение, то есть они образованы при участии сразу несколько зародышевых листков. Например, мускулатура пищеварительного тракта является производным мезодермы, а его внутренняя выстилка — производное энтодермы. Однако, несколько упрощая, происхождение основных органов и их систем все-таки можно связать с определенными зародышевыми листками.
| Нейруляция |
Зародыш на стадии нейруляции называется нейрулой (рис. 317). Материал, используемый на построение нервной системы у позвоночных животных — нейроэктодерма, входит в состав спинной (дорсальной) части эктодермы. Он располагается над за
|
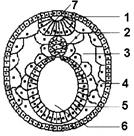 Рис. 317. Нейрула:
1 — эктодерма; 2 — хорда; 3 — вторичная полость тела; 4 — мезодерма; 5 — энтодерма; 6 — кишечная полость; 7 — нервная трубка.
Рис. 317. Нейрула:
1 — эктодерма; 2 — хорда; 3 — вторичная полость тела; 4 — мезодерма; 5 — энтодерма; 6 — кишечная полость; 7 — нервная трубка.
является одним из наиболее важных во всем развитии. Сначала в области нейроэктодермы происходит уплощение клеточного пласта, что приводит к образованию нервной пластинки. Затем края нервной пластинки утолщаются и приподнимаются, образуя нервные валики. В центре пластинки за счет перемещения клеток по средней линии возникает нервный желобок, разделяющий зародыш на будущие правую и левую половины. Нервная пластинка начинает складываться по средней линии. Края ее соприкасаются, а затем смыкаются. В результате этих процессов возникает нервная трубка с полостью — невроцелем.
Смыкание валиков происходит сначала в средней, а затем в задней части нервного желобка. В последнюю очередь это происходит в головной части, которая по ширине превосходит другие. Передний, расширенный отдел в дальнейшем образует головной мозг, остальная часть нервной трубки — спинной. В результате нервная пластинка превращается в нервную трубку, лежащую под эктодермой.
В ходе нейруляции часть клеток нервной пластинки не входят в состав нервной трубки. Они образуют ганглиозную пластинку, или нервный гребень, — скопление клеток вдоль нервной трубки. Позднее эти клетки мигрируют по всему зародышу, образуя клетки нервных узлов, мозгового вещества надпочечников, пигментные клетки и т.п.
| Образование систем органов |
Из материала эктодермы, помимо нервной трубки, развиваются эпидермис и его производные (перо, волосы, ногти, когти, кожные железы и т.д.), компоненты органов зрения, слуха, обоняния, эпителий ротовой полости, эмаль зубов.
Мезодермальные и энтодермальные органы формируются не после образования нервной трубки, а одновременно с ней. Практически одновременно с нейруляцией происходят процессы закладки мезодермы и хорды. Вначале вдоль боковых стенок первичной кишки путем выпячивания энтодермы образуются карманы, или складки. Участок энтодермы, расположенный между этими складками, утолщается, прогибается, сворачивается и отшнуровывается от основной массы энтодермы. Так появляется хорда. Возникшие карманообразные выпячивания энтодермы отшнуровываются от первичной кишки и превращаются в ряд сегментарно-расположенных замкнутых мешков, называемых также целомическими мешками. Их стенки образованы мезодермой, а полость внутри представляет собой вторичную полость тела (или целом).
Из мезодермы развиваются все виды соединительной ткани, дерма, скелет, поперечно-полосатая и гладкая мускулатура, кровеносная и лимфатическая системы, половая система.
Из материала энтодермы развивается эпителий кишечника и желудка, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, секретирующие отделы передней и средней доли гипофиза, щитовидной и паращитовидной желез.
| Эмбриональная индукция |
Наблюдения за оплодотворенной яйцеклеткой лягушки позволили проследить путь развития клеток, входящих в состав того или иного участка зародыша. Оказалось, что определенные клетки, занимающие соответствующее место в бластуле, дают начало строго определенным зачаткам органов. Удалось выяснить, какие группы клеток дают начало нервной трубке, хорде, мезодерме, кожному эпителию и т.д. Действительно, в развивающемся организме различные группы клеток дают начало определенным органам и тканям, а культивирование клеток вне зародыша (в пробирке) не приводит к формированию типичных тканевых структур, которые должны были бы образоваться из клеток. Чем же вызывается преобразование тех или иных клеток зародыша в конкретные ткани и органы?
В 1924 г. были опубликованы результаты опытов Г.Шпемана и Г.Мангольда, посвященные выяснению этого вопроса (рис. 318). На стадии ранней гаструлы зачаток эктодермы, который в нормальных условиях должен был развиться в структуры нервной системы, из зародыша гребенчатого (непигментриованного) тритона пересаживался под эктодерму брюшной стороны, дающую начало эпидермису кожи, зародыша обыкновенного (пигментированного) тритона. В итоге на брюшной стороне зародыша-реципиента возникала сначала нервная трубка и другие компоненты комплекса осевых органов, а затем формировался дополнительный зародыш. Причем, наблюдения показали, что ткани дополнительного зародыша формируются почти исключительно из клеточного материала реципиента.
Эти данные доказывают, что в ходе эмбриогенеза некоторые части зародыша влияют на пути развития соседних участков. Такое влияние одного зачатка на другой получило название эмбриональной индукции. Насколько важную роль играет эмбриональная индукция в развитии, показывает следующий опыт. Если на стадии ранней гаструлы полностью удалить
|
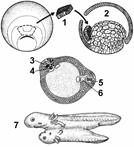 Рис 318. Эмбриональная индукция:
1 — зачаток хордомезодермы; 2 — полость бластулы; 3 — индуцирорванная нервная трубка; 4 — индуцированная хорда; 5 — первичная нервная трубка; 6 — первичная хорда; 7 — формирование вторичного зародыша, соединенного с зародышем-хозяином.
Рис 318. Эмбриональная индукция:
1 — зачаток хордомезодермы; 2 — полость бластулы; 3 — индуцирорванная нервная трубка; 4 — индуцированная хорда; 5 — первичная нервная трубка; 6 — первичная хорда; 7 — формирование вторичного зародыша, соединенного с зародышем-хозяином.
При дальнейшем изучении развития зародышей оказалось, что зачаток хордомезодермы представляет собой не только индуктор нервной трубки, но и сам для дифференцировки нуждается в индуцирующем влиянии со стороны зачатка нервной системы. Во время эмбрионального развития имеет место не односторонняя индукция, а взаимодействие частей развивающегося зародыша. Таким образом, эмбриональную индукцию можно определить как явление, при котором в процессе эмбриогенеза один зачаток влияет на другой, определяя путь его развития, и, кроме того, сам подвергается индуцирующему воздействию со стороны первого зачатка.
|
|
|
|
|
Дата добавления: 2015-06-27; Просмотров: 785; Нарушение авторских прав?; Мы поможем в написании вашей работы!