КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Б. Показатели биологической продуктивности
|
|
|
|
Количественное выражение вещественно-энергетических потоков может служить основой для оценки эффективности продукционных процессов и средообразующих функций ценоза. Для этой цели разработано множество показателей.
Ниже дается трактовка основных терминов, используемые при рассмотрении изменений биомассы и биологической продукции. Различают общую, первичную и вторичную продукцию.
1. Общая продукция (биомасса) – запасы всей органической массы, т.е. масса организмов, присутствующих в экосистеме в момент наблюдения и учтенная на единицу площади. Легче всего поддается учету и измерению запасы биомассы и продукция живых растений – автотрофов, она называется фитомассой. Зоомассой, соответственно, называется масса консументов.
Все "массы" могут быть выражены в единицах массы и энергии (калориях).
2. Первичной продукцией, или брутто-продукцией, называется органическая масса, создаваемая продуцентами в единицу времени. Первичная валовая продукция (Пв) — суммарная продукция фотосинтеза (суммарная ассимиляция), включающая, следовательно, и вещество, сжигаемое при дыхании за единицу времени.
Чистая первичная продукция (Пч), или нетто-продукция, — это суммарная продукция фотосинтеза за вычетом вещества, затраченного растениями на дыхание, т.е. вещество, которое можно взвесить при уборке урожая (табл.). Определяется эффективностью фотосинтеза, площади фотосинтезирующих органов, а также от длительности периода активного фотосинтеза. Для многолетних растений Пч – масса годичного прироста.
Таблица
Значения чистой первичной (годовой) продукции некоторых культур (растения целиком), т/га абс.-сух вещества
| Культура | Чистая продукция (Пч) | Страна | |
| всего в мире | по стране | ||
| Пшеница | 3,44 | 12,50 | Голландия |
| Овес | 3,59 | 9,26 | Дания |
| Кукуруза | 4,12 | 7,90 | Канада |
| Рис | 4,97 | 14,40 | Япония, Италия |
| Травы | 4,20 | 9,40 | США (Калифорния) |
| Картофель | 3,85 | 8,45 | Голландия |
| Сахарная свекла | 7,65 | 14,70 | Голландия |
| Сахарный тростник | 17,25 | 34,30 | США (Гавайи) |
Согласно Ю. Одуму (1959), значения первичной продукции для разных типов биогеоценозов следующие:
1. Очень низкой продуктивностью обладают океанические глубины и пустыни (0,1 г/м2 в день). Хотя ограничивающие факторы двух сред совершенно различны (в океанах — питательные вещества, в пустынях — вода), как различны и продуцирующие организмы (водоросли — в океанах), глубины океанов и аридные территории по продуктивности можно рассматривать как пустыни.
2. Сходной низкой продуктивностью (0,5 — 3 г/м2 в день) характеризуются травяные формации, морские литорали и плохо культивированные земли.
3. Продуктивностью от 10 до 20 г/м2 в день обладают влажные леса, земли с интенсивным сельскохозяйственным использованием, аллювиальные формации, эстуарии и коралловые рифы. Максимально возможная продуктивность экосистемы, по-видимому, не может превышать 25 г/м2 в день.
3. Вторичная продукция (П2) – прирост за единицу времени массы консументов.
4. Опад – вся отмершая масса растений, лежащая на поверхности – мелкие растительные остатки, характеризующиеся поэтому более быстрыми в обычных условиях темпами разложения (листву, генеративные органы многих растений, ветви и побеги толщиной менее 5 мм, травы, мхи и лишайники, кусочки коры и др.)
5. Отпад – отмершие органы растений – более крупные, чем опад. Обычно сильно лигнизированные остатки (стволы, ветви толщиной более 5 мм, шишки хвойных пород, крупные куски коры деревьев).
6. Величина годичного опада и отпада органического вещества растительного происхождения, определяемая по массе и энергии и с учетом ярусного сложения фитоценозов.
7. Годичная норма разложения растительных остатков как результат деятельности сапрофагов и редуцентов и физико-химических процессов в слое лесной подстилки.
8. Фракционный состав запаса и годичного прироста фитомассы; выражается процентным соотношением массы разных фракций фитомассы: стволы, ветви и побеги, листва, генеративные органы, корни. В пределах этих групп фракций желательна дальнейшая дифференциация: для стволов, ветвей и корней – на древесину и кору; для ветвей и корней – по группам толщины; для хвои вечнозеленых пород – по возрастам; для генеративных органов с целью полного учета общего годичного прироста – на цветки (или стробилы); недоразвившиеся плоды (или женские стробилы) – на семена и другие части плодов (или женских стробил) в зрелом состоянии
9. Соотношение массы надземных частей растений к массе корней, определяемое: а) в запасе; б) в годичном приросте фитомассы.
10. Отношение суммарной длины древесных частей растений в надземном слое к суммарной длине корней, если последний показатель будет определен.
11. Коэффициенты связи между весом разных частей деревьев и таксационными признаками стволов. Эти показатели, в зависимости от принятого метода обработки данных модельных деревьев, могут быть представлены либо значениями постоянных коэффициентов аллометрических уравнений (регрессий), либо средними значениями относительных коэффициентов, получаемых при делении веса разных частей деревьев на d2 или на d2h (Ватковский, 1968), где d – диаметр ствола на высоте груди (в см), h – высота ствола (в м).
12. Коэффициент продуктивности (или эффективности работы) ассимиляционного аппарата растений, выражается отношением общего годичного прироста (нетто-продукции) к массе листвы.
13. Индекс листовой поверхности LAI (общий), или отношение площади поверхности листвы (для листьев берется односторонняя поверхность, для хвои – общая) отдельных ярусов и фитоценозов в целом к площади участка в 1 га..
14. Коэффициент эффективности работы трофического уровня продуцентов; определяется по отношению энергетических эквивалентов для общего и чистого годичных приростов фитомассы к количеству солнечной энергии, поступающей в систему в течение года и (отдельные расчеты) за вегетационный период.
15. Отношение массы листвы к массе всасывающих корней (в расчете на 1 га). Перечисленные и многие другие продукционные показатели отражают разные стороны жизнедеятельности биогеоценоза, его географические, экотопические и возрастные особенности, взаимоотношения между компонентами в борьбе за жизненные факторы (влагу, тепло, свет). Во многом они сходны, в первую очередь сходны тенденции изменения разных показателей, в то же время могут существенно различаться. Так, в зависимости от указанных выше факторов, в лесных биогеоценозах сильно варьирует структура древостоя. Доля стволовой массы может колебаться от 40-50 до 80-85 %. В условиях тайги приближенная доля средних значений участия стволовой древесины в фитомассе древостоя колеблется примерно от 45 до 60 %; доля крон – от 12 до 35 %; доля корней – от 11 до 22 %.
Ниже, в качестве примера, приводятся основные результаты анализа первичной продуктивности, изученной нами в Северном Охотоморье (юг Магаданской области).
I. Характеристика исследованных фитоценозов
Северное побережье Охотского моря характеризуется преобладанием горных форм рельефа и суровым климатом, обусловливающим длительное сохранение сезонной и локальное распространение многолетней мерзлоты в почвах. В горах хорошо выражена высотная дифференциация растительного покрова, представленная горными тундрами (на отметках выше 400 м над ур. м.), поясом кедровостланиковых зарослей (400-200 м), лиственничными рединами и редколесьями с подлеском из кедрового стланика. Высокополнотные лиственничники и тополево-чозениевые леса занимают лишь речные поймы.
Исследования по биологической продуктивности выполнены в топоэкологическом ряду из пяти фитоценозов, представляющих основные типы леса юга Магаданской области, в том числе четыре лиственничных (разнотравно-хвощовый, зеленомошно-брусничный с подлеском из кедрового стланика, бруснично-лишайниковый с подлеском из кедрового стлaника и осоково-сфагновый) и один лиственный - чозенник с тополем разнотравно- хвощовый.
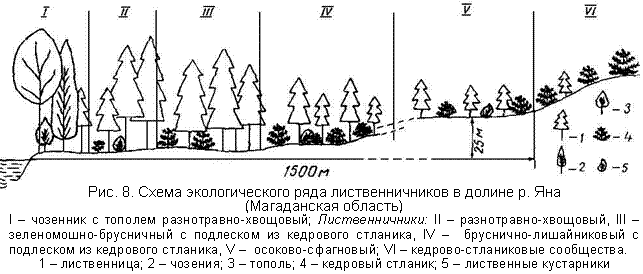
| Каждый из изученных типов леса занимает определенный геоморфологический уровень в пределах речной долины р. Яна (рис. 8) и сформирован при определенном сочетании эдафических факторов. Так, прирусловый чозенник с тополем разнотравно-хвощовый произрастает в условиях регулярного затопления паводками и неглубокого зеркала подрусловых вод, отепляющих слаборазвитые, но достаточно плодородные аллювиальные почвы. |
Лuственнuчнuк разнотравно-хвощовый занимает высокие участки речной поймы с аллювиальными дерновыми почвами, редко затопляющиеся паводковыми водами (в среднем один раз в 10 лет).
Лиственнuчник зеленомошно-6русничный с подлеском из кедрового стланика приурочен к первой речной террасе со старопойменными глееватыми суглинистыми почвами, сравнительно недавно вышедшими из-под влияния пойменного режима и лишенными отепляющего воздействия подрусловых вод. Они характеризуются значительным содержанием токсичного для растений закисного железа и периодическим переувлажнением.
Лиственничник 6руснично-лишайниковый с подлеском из кедрового стланика произрастает на подзолах Al-Fе-гумусовых второй речной террасы. Эти почвы летом прогреваются значительно сильней почв других типов леса, но характеризуются сухостью, высокой кислотностью и низким содержанием элементов питания.
Лuственнuчнuк осоково-сфагновый развивается на склоне коренного берега с болотными мерзлотными почвами, имеющими крайне неблагоприятные водно-физические и физико-химические свойства. Для них характерны постоянное переувлажнение, наличие близко залегающей многолетней мерзлоты, бедность питательными веществами, сильнокислая реакция и др.
II. Запасы и фракционная структура биомассы
Анализ представленных материалов позволил установить:
1. Особенности накопления и структуры растительной массы в лесах Северного Охотоморья позволяют отнести их к типичным северотаежным сообществам. Их фитоценозы обладают сходной фитоценотической структурой и продуктивностью с фитоценозами других районов северотаежной подзоны. В них к возрасту биологической спелости древостоев накапливается 55-125 т/га растительной массы и лишь в наиболее продуктивных - до 310 т/га; масса годичного прироста составляет соответственно 2,2-7,8 т/га.
2. Наиболее сложным строением и максимальной продуктивностью (до 310 т/га) характеризуются разнотравно-хвощовые типы леса, произрастающие на разных уровнях поймы. С выходом насаждений из зоны влияния пойменного режима и прекращением отепляющего воздействия подрусловых вод, ухудшением гидротермических свойств почв, снижением в них содержания доступных растениям элементов питания, усилением кислой реакции среды и т. д. заметно упрощается их фитоценотическая структура, обедняется видовой состав, сокращается объемное пространство, занятое фитоценозом; общие запасы и годичный прирост биомассы уменьшаются в 2,5-5,6 раза.
3. В исследованных насаждениях масса годичного прироста варьирует от 2,2 до 7,7 т/га; максимальное значение - в лиственничнике разнотравно-хвощовом, минимальное – в осоково-сфагновом. Наиболее эффективным использованием солнечной энергии характеризуется тополево-чозениевый фитоценоз, в древесине которого откладывается до 46% массы годичного прироста, тогда как в лиственничниках - не более 35%.
4. Коэффициент эффективности работы ассимилирующих органов в чозеннике с тополем равен 1,89 и снижается до 0,33 в осоково-сфагновом лиственничнике.
5. Менее продуктивным фитоценозам соответствует более высокое содержание отмершей подземной массы. В растительной массе лиственничников на ее долю приходится 4 (разнотравно-хвощовый) – 24% (осоково-сфагновый). Накопление на поверхности и в почве большого количества отмерших растительных остатков обусловлено неблагоприятным гидротермическим режимом почв, их кислой реакцией, высоким содержанием в растительных остатках восков и смол, малочисленностью и малой активностью почвенной микрофлоры.
6. Во всех типах леса основные запасы растительной массы насаждений образованы древостоем – 66 (осоково-сфагновый) – 99% (разнотравно-хвощовые). Максимальные запасы их характерны для разнотравно-хвощовых типов леса. В низко продуктивных насаждениях они снижаются почти в 9 раз, а масса напочвенного покрова увеличивается в 16 раз.
Тополево-чозениевый фитоценоз имеет сходные с высокопродуктивными лиственничниками запасы биомассы древостоев, но отличается от них более высоким содержанием надземных аккумулирующих органов.
7. В массе растущих деревьев на долю надземных органов приходится 47-76%, стволов - 39-70%; меньшие значения соответствуют менее продуктивными фитоценозам.
8. С ухудшением экологических условий происходит перераспределение биомассы в пользу ассимилирующих органов, т.е. тех, которые в первую очередь обеспечивают выполнение жизненно важных функций. Это фотосинтезирующий аппарат и мелкие корни. В исследованных лиственничниках хвоя составляет 13-17% кроновой и 1,4-2,9% надземной массы, корни 34-53% общей массы растущих деревьев. Менее продуктивным фитоценозам соответствуют большие значения этих показателей.
Участие корней в формировании растительной массы рассмотренных древостоев в 1,5-2,0 раза превышает известные в литературе данные для более западных районов северотаежной подзоны, что, очевидно, вызвано более суровыми климатическими условиями на Дальнем Востоке.
9. Первичный экотоп не всегда является определяющим в формировании того или иного компонента фитоценоза, поскольку на действие абиотических факторов среды накладывается лимитирующее влияние эдификатора сообщества. Оптимальныe условия для произрастания кедрового стланика под пологом леса свойственны не разнотравно-хвощовым типам леса, в которых средообразующее влияние лиственницы особенно высоко, а зеленомошно-брусничному лиственничнику. В этом насаждении стланик характеризуется наименьшим содержанием корней и самой высокой продуктивностью хвои - отношение массы многолетних надземных органов к массе хвои равно 6:1, тогда как в других исследованных фитоценозах оно составляет 4,5:1-2,7:1.
10. В растительной массе исследованных насаждений отмершая подземная составляет 3 (чозенник с тополем) – 24% (осоково-сфагновый). Преимущественное накопление ее в низкобонитетных насаждениях вызвано несоответствием между относительно большим количеством ежегодных растительных опадов и крайне низкими темпами их разложения.
12. Нижние ярусы образуют от 1 до 35% общего запаса и от 1 до 17% фитомассы насаждений. Увеличение запасов, особенно напочвенного покрова, происходит по мере обеднения и ухудшения гидротермического режима почв и упрощения морфоструктуры фитоценозов. При этом резко сокращаются запасы лиственных кустарников и трав и увеличиваются запасы мхов при избыточном и лишайников при недостаточном увлажнении.
13. Коэффициент эффективности работы ассимилирующих органов в чозеннике с тополем равен 1,89 и снижается до 0,33 в осоково-сфагновом лиственничнике, в котором вследствие полной деградации древесно-кустарниковой растительности вся поверхность представлена сплошным, внешне однородным, осоково-сфагновым покровом.
III. Вертикальный профиль кроновой и зеленой массы

| 1. Распределение по высоте фитомассы крон древостоя и подлеска и фото-синтезирующих органов подтверждает правильность вычленения в исследованных типах леса вертикальных структурных элементов – фитогоризонтов. Оно позволяет объективно оценить их функциональную роль. Выявлено, что чем выше средообразующее значение древесного или кустарникового яруса, тем большим запасом кроновой и зеленой массы в однометровом слое он обладает. 2. Степень эдификаторных функций лиственничных древостоев отражается в вертикальной структуре кроновой и зеленой массы: чем сильнее они проявляются, тем отчетливее выражена концентрация массы этих фракций в верхних слоях кронового полога (рис. 10). 3. Высокополнотные древостои отличаются неравномерным охвоением кронового полога по высоте. Наибольшим содержанием хвои отличаются слои, где наблюдается перекрытие крон смежных ярусов, т.е. световая обстановка характеризуется напряженностью. 4. В редкостойных лиственничниках в связи с благоприятными радиационными условиями по всему кроновому пологу отмечено одинаковое охвоение крон на протяжении всего вертикального профиля и только в осоково-сфагновом лиственничнике оно резко увеличивается (с 17 до 30%) в верхних слоях. По-видимому, древостои однозначно реагируют не только на ухудшение светового режима, но и на ограничения других факторов, действующих в надземном пространстве. В данном случае наиболее вероятным является недостаточная теплообеспеченность верхней части полога фитоценоза. |
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 1463; Нарушение авторских прав?; Мы поможем в написании вашей работы!