КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Метилирование ДНК и сайленсинг у растений
|
|
|
|
Для растений характерно:
1) Разветвленная система метилирования: cайты метилирования - CpG, а также CpHpG и CHH (H=T, C или А)
2) Мутации по системам метилирования нелетальны, но ограничивают рост и нарушают морфогенез, что позволяет проводить генетический анализ этих систем
3) Метилированы центромерные районы, рДНК повторы, транспозоны
4) Сайленсинг транспозонов у растений обеспечивается
- CpG метилированием при репликации, не зависящим от siРНК
- CpHpG и CHH метилированием, зависящим от siRNA и AGO4 (pol IV и pol V)
РНК-зависимое метилирование ДНК растений:
1) CрG метилирование при репликации mCpG mCpG
GpC GpCm
2) СНG и СНН метилирование, где Н= A, C или Т (non-CpG); РНК-зависимое метилирование RNA dependent methylation, RdM.
Уникально то, что можно делать и мутации и смотреть их эффект (они не летальны). Метилирование ДНК и сайленсинг у растений уникальная область. Метилирование у растений напоминает piРНК дрозофилл.
Эпигенетический ландшафт Arabidopsis.
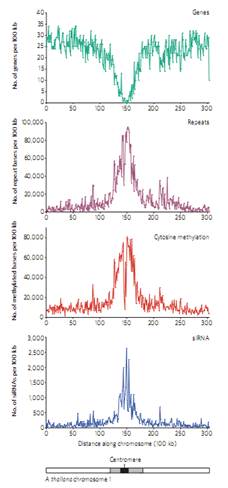 По оси ординат количество генов на единицу длины ДНК, по оси абсцисс – kb.
По оси ординат количество генов на единицу длины ДНК, по оси абсцисс – kb.
Это хромосома 1, она схематически обозначена внизу серой полоской. Около центромеры гетерохроматин.
Видно, что в районе центромеры практически нет белок-кодирующих генов, зато много повторов, метилированных по 5-ому положению цитозинов, а также siРНК. Где повторы, там идет в значительной степени метилирование. Это нужно для того, чтобы запретить здесь рекомбинацию.
Можно секвенировать siРНК, которая образуется в растениях, и эти последовательности накладывать на геномные последовательности, и оказывается, что все siРНК попадают в район, где идет метилирование 5mC. Эти данные показывают, что метилирование связано с siРНК.
Как это все происходит. Здесь играет роль две новые РНК-полимеразы: полимераза IV и V. Похожи на полимеразу II, поскольку содержат общие субъединицы, являются специализированными РНК-полимеразами растений, синтезирующие нкРНК. Полимераза IV (первый этап) направлена на синтез siРНК, другая полимераза V (второй этап) - метилирование собственно ДНК. Не все siРНК возникают таким путем. Целый ряд siРНК в результате перекрывающейся транскрипции (полимераза II) (как было сказано ранее), а если имеются шпильки при транскрипции, то они могут складываться и давать двуцепочечные РНК, которые затем разрезаются дайсером.
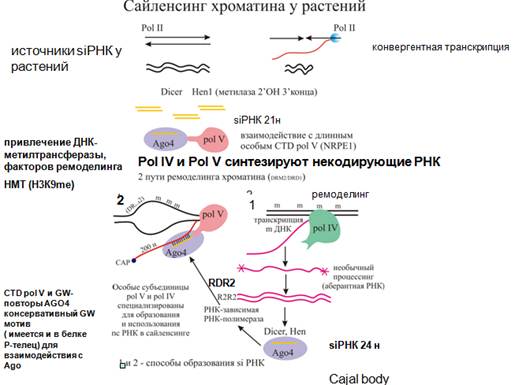
Что известно о полимеразах IV и V: обе эти полимеразы и синтезируют довольно плохо. In vitro ничего не синтезируют. Для полимеразы IV оказывается доступной даже метилированная ДНК. Полимераза IV начинает ее синтезировать. И полимераза IV, и соответствующие транскрипты обладают таким свойством, что сюда к транскриптам привлекается РНК-зависимая-РНК- полимераза, которая делает дцРНК, и дальше соответствующий дайсер делает siРНК 24 нуклеотидов длиной. Еще имеется фермент, который на 3' конец вешает метильную группу, с чем мы уже сталкивались, когда рассматривали siРНК. Образовавшиеся siРНК связываются со специфическим Аргонавтом. Комплекс Аргонавта с siРНК прочитывается полимеразой V в результате наличия комплементарной последовательности (комплементарность между siРНК и транскриптом) и далее осуществляется метилирование ДНК. Для чего все это представляется нужно. Короткая РНК образовалась, для того чтобы гомологичные нуклеотидные последовательности заметилировать в геноме. Фактически, это транскрипция мобильных элементов и других повторов полимеразой IV. Поэтому накапливается большое количество siРНК, которые потом уже используются здесь для того, чтобы осуществить метилирование. Показано, что большая субъединица полимеразы V имеет такой домен, который взаимодействует с определенными доменами Аргонавта 4 (Аgo4). За счет этого обеспечивается связывание и приманивание siРНК сюда и привлечение комплексов, которые ремоделируют хроматин для того, чтобы ДНК-метилаза распознала мишень и осуществила метилирование. Процессы идут у растений тоже в специализированных ядерных тельцах Кахаля, которые есть у растений и животных.
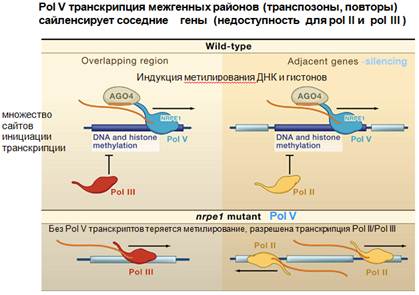
Полимераза V транскрибирует межгенный район, а рядом находится белок-кодирующих ген. И благодаря тому, что полимераза V осуществляет метилирование соответствующей мишени которую она транскрибирует, осуществляется распространение гетерохроматина, который образовался в виде метилированной ДНК, на соседние участки, которые больше не могут транскрибироваться полимеразой II. В тех случаях, когда мы имеем мутацию в полимеразе V, то тогда соответствующие участки уже доступны для транскрипции полимеразой III или для транскрипции полимеразой II. Когда нет соответствующей транскрипции, нет метилирования с участием полимеразы V, то разрешается транскрипция обычной полимеразой II.
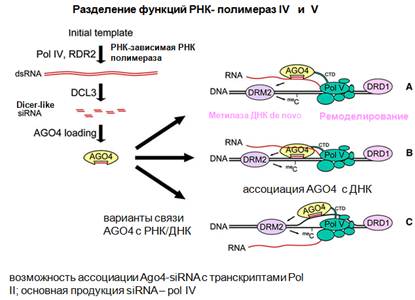
На картинке показано хорошо разделение функций двух полимераз V и IV. Есть некая изначальная матрица, шаблон, по которому идет транскрипция, в основном это повторяющиеся элементы. Образуется дцРНК, образуются короткие РНК, образуется комплекс Аргонавт с ними. Далее, что происходит с транскриптами, которые должны метилироваться. Комплекс с Аргонавтом взаимодействует с растущей РНК, либо с новообразованной РНК, либо с ДНК. Либо он может взаимодействовать с одной нитью ДНК, есть разные возможности, но наиболее вероятно с растущей цепью РНК. У растений есть несколько ДНК-метилаз, одни из них поддерживают метилирование. Когда осуществляется репликация, одна нить метилирована, другая нет, и ДНК-метилаза узнает такое состояние и метилирует комплементарную нить. Это так называемое поддерживающие метилирование. А есть метилазы, которые привлекают комплексы ремоделирования (фиолетовым овалом обозначено). Надо сказать, что не исключена возможность ассоциации этого комплекса с Аргонавтом и siРНК и транскрипции полимеразой II. Здесь тоже может за счет транскрипции осуществляться сайленсинг. Ремоделирование поддерживается при делении клеток и наследуется эпигеномно.
Теперь рассмотрим пример. Возникновение парамутаций.
Парамутация – это перенос приобретенного неактивного эпигеномного состояния на гомологичный локус в другой хромосоме (кукуруза, мышь). Свойства:
• Наследуемость этого состояния в следующих поколениях
• Эффект парамутации зависит от РНК-зависимой РНК полимеразы и полимеразы IV, транскрибирующей метилированные последовательности, процессируемые в siРНК
• Свидетельство о существовании siРНК зависимой эпигеномной модификации
Эта необычная мутация. Заключается в следующем: если есть некое мутантное неактивное состояние гена, и после скрещивания в гомологичной хромосоме напротив этого мутантного гена имеется нормальный аллель, то он становится тоже мутантным, то есть осуществляется перенос неактивных эпигеномных состояний на гомологичный локус в другой хромосоме. Впервые был показан на кукурузе. По-видимому, это достаточно общее явление, поскольку недавно подобное было показано у дрозофилы, у млекопитающих. Это приобретенное состояние гомологичных хромосом может наследоваться в нескольких поколениях.
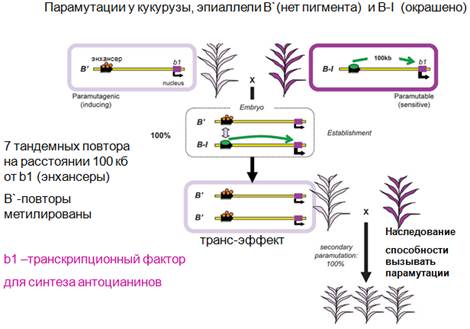
У кукурузы генетика изучена достаточно хорошо. Как мы уже видели, этапы участия полимеразы V и IV были исследованы на арабидопсис. А парамутации будут рассмотрены на кукурузе. Было показано, что возникновение парамутации зависит активности генов, которые кодируют РНК-зависимую-РНК-полимеразу и полимеразу IV. Возникновение парамутации свидетельствует о том, что существует некая эпигеномная наследуемая модификация с участием siРНК. Теперь на конкретном примере, классическая форма парамутации. Речь пойдет о локусе В-1, который определяет фактор транскрипции, отвечающий за синтез антоцианинов. На расстояние от этого локуса около 100 kb находится энхансер, который представлен повтором. Существует активное состояние этого локуса (справа). Зелеными стрелками показано расстояние от энхансера до белок-кодирующего локуса. Слева локус плохо работает. Среди аллелей есть эпиаллель В1, и есть В', который является малоактивным. Когда локус активен, растение окрашено, в другом случае (локус малоактивен) - не окрашено. Осуществляется скрещивание двух растений. Получается гетерозиготное состояние. Показано, что энхансер влияет на соответствующий ген и обеспечивает образование транскрипционного фактора. В гетерозиготном состоянии осуществляется транс-эффект, он заключается в том, что активный эпиаллель превращаются в неактивный, и в потомстве такого растения окраски не видим. То есть никакой доминантности активного аллеля нет, и более того, неактивное состояние, способность вызывать парамутации, делать локус мутагенным, наследуется. Если дальше скрещивать, то будут получаться растения не обладающие пигментом.
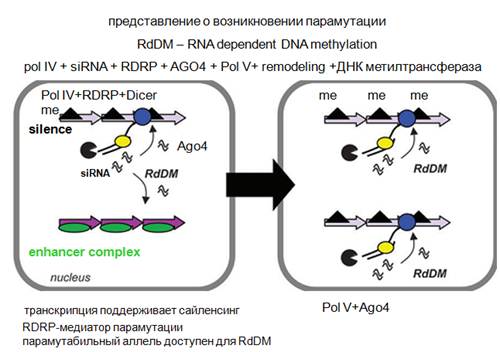
Детально механизм возникновения парамутации не изучен. Чисто генетический анализ показывает, что для того, чтобы все это происходило (возникновение парамутагенного состояния), нужна полимераза IV, а также должна образоваться siРНК, должна быть РНК-зависимая-РНК-полимераза, Аргонавт, факторы ремоделирования хроматина, ДНК-метилтрансфераза. Здесь показано неактивное состояние, где осуществляется метилирование повторов энхансера. Энхансер состоит из отдельных повторов. Полимераза IV может все читать, РНК-зависимая-РНК-полимераза делает комплементарную матрицу, дайсер это нарезает на кусочки, далее этот активный энхансерный комплекс, состоящий из повторов превращается в неактивный комплекс за счет того, что начинает метилироваться. Если эти повторы будет транскрибировать полимераза V (картинка справа), наработанные здесь siРНК приведут к метилированию этого района. Детально это не изучено (биохимически).
Посмотрим, как ведет себя siРНК в генеративных клетках: в пестиках, тычинках, рассмотрим участие в оплодотворении и роль клеток «компаньонов», которые сопровождают клетки спермы или соседствует с яйцеклеткой. Имеется аналогия с функционированием piРНК у животных. Мы увидим, что в генеративных клетках и клетках-«компаньонах» может осуществляться направленная активация транспозонов. В зрелой пыльце имеется 1 крупная вегетативная клетка с вегетативным ядром и две более мелкие клетки, которые называются sperm cells (ядра называют - ядра спермы растений – он так сказал).
|
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 1816; Нарушение авторских прав?; Мы поможем в написании вашей работы!